- Published:
Crinoids from the Middle Jurassic (Bajocian–Lower Callovian) of Ardèche, France
Swiss Journal of Palaeontology volume 131, pages 211–253 (2012)
Abstract
Several Middle Jurassic outcrops in the Ardèche Department near La Voulte-sur-Rhône and St-Étienne-de-Boulogne are rich in the remains of crinoids, but these were known from surface collections only and were not described using present standards of systematics. This paper brings the taxonomic status of the previously described crinoids up to date, reassesses the systematic position of some of the species based on cups and describes new forms. Sampling and washing of bulk material from the Lower Bathonian of the La Pouza locality yielded nearly 100,000 crinoid ossicles. Among them are rare comatulids with the following recognized as new: Andymetra galei n. g., n. sp., Palaeocomaster messingi n. sp., Singillatimetra inordinata n. g., n. sp. and Solanocrinites voultensis n. sp. These forms supplement the meagre record of Middle Jurassic comatulids and indicate that this group radiated well before the Late Jurassic. The Cyrtocrinida constitute the bulk of the crinoids and they are dominated by Cyrtocrinus praenutans n. sp. from which nearly all parts of the skeleton are described in their morphological variability. The La Pouza site furnished the additional new cyrtocrinids, Praetetracrinus bathonicus n. sp. and Phyllocrinus voultensis n. sp. The material also includes numerous remains of the isocrinids Isocrinus dumortieri (de Loriol), Balanocrinus dumortieri de Loriol, Balanocrinus pacomei de Loriol and Balanocrinus inornatus (d’Orbigny), which are described in some detail, including brachials. From the Upper Bajocian–Lower Bathonian locality of Pont des Étoiles, Pentacrinites ausichi n. sp. and the cyrtocrinid Scutellacrinus tenuis n. g., n. sp. are new to science. The results demonstrate that the Middle Jurassic crinoids from the Ardèche are one of the important and diverse Mesozoic crinoid faunas. Some forms bridge the gap between the Early Jurassic and the Late Jurassic hardground faunas of cyrtocrinids. Cyrtocrinus praenutans n. sp., a form similar to Cyrtocrinus nutans (Goldfuss) from the Oxfordian, is described as a separate species despite some overlapping phenotypic variability of cups and columnals. Pathological deformations on all types of ossicles of C. praenutans n. sp. are ascribed to the epizoan commensal Oichnus paraboloides Bromley. Different species are dominant at the different Bathonian localities, namely C. praenutans n. sp. at La Pouza and Phyllocrinus fenestratus (Dumortier) and Lonchocrinus dumortieri (de Loriol) at La Clapouze. Preservation and rock formation of the Upper Bajocian–Lower Bathonian Isocrinus nicoleti (Thurmann) at Pont des Étoiles suggests that this form lived in rather shallow and turbulent water. The dominance of cyrtocrinids and the presence of all growth stages and parts of the skeleton at La Pouza and La Clapouze suggest a deeper palaeoenvironment, with some transport of the ossicles.
Introduction
In the Ardèche department, several Middle Jurassic outcrops have yielded rich assemblages of crinoids, especially cyrtocrinids. After the Early Jurassic radiation (Hess 2006), this was the second major radiation of this important and intriguing group of Mesozoic–Holocene crinoids, which dominated hardground faunas from Early Jurassic to Early Cretaceous times, reaching their apogee in the Valanginian of Štramberk (Czech Republic), with approximately 50 taxa (see Ausich et al. 1999, p. 48). The crinoids from the Middle Jurassic of Ardèche were described by Dumortier (1871), de Loriol (1882) and, recently, by Charbonnier et al. (2007). However, one of the localities was never included and material was from surface collections only. The present paper is based on material from the author’s field collections in the late 1950s, when the outcrops in question furnished rich material (and the author’s eyes were better). Two cyrtocrinid cups from the Gevrey collection of the Université Joseph Fournier of Grenoble and labelled “Bathonien, Rompon” are also included in the present study. The most common cyrtocrinid from the localities near La Voulte-sur-Rhône has been ascribed to Cyrtocrinus nutans (Goldfuss). This form is also common in the Middle Oxfordian sponge facies of Switzerland and Germany. It is intriguing that a distinct cyrtocrinid species should range from the Upper Bajocian to the Middle Oxfordian, a period of some 15 m.a. Because such longevity seems exceptional for a stalked crinoid, the variability of specimens from the Middle and Late Jurassic occurrences are discussed in some detail.
Recently, A. S. Gale sampled approximately 100 kg bulk sediment from three Bathonian beds near the La Pouza farmhouse. Subsequent processing has yielded nearly 100,000 crinoid ossicles that are included in the present study. These remains furnish information on nearly the whole range of ossicles and all developmental stages. The rich material also includes some rare comatulids, not normally found by surface collecting. The present paper updates the taxonomic status of the previously described crinoids, reassesses the systematic position of some of the species based on cups, and describes new forms. The results demonstrate that the Middle Jurassic crinoids from the Ardèche are one of the important Mesozoic crinoid faunas.
Geological setting and fossil localities
The area in question is part of the eastern sedimentary rock cover of the Massif Central, a Hercynian crystalline complex whose eastern margin was faulted and tilted during the Middle Jurassic; it was situated along the western part of the Tethys, with a complex submarine topography of platforms, escarpments and basins (Alméras & Elmi 1996). Near La Voulte-sur-Rhône are crinoid-rich outcrops of Upper Bajocian to Lower Callovian age (Fig. 1). They include the locality Pont des Étoiles with Upper Bajocian/Lower Bathonian crinoidal limestones and marls, the locality La Pouza with Lower Bathonian marls and marly limestones containing siliceous sponges and brachiopods, and the Lower Callovian locality of Chénier Ravine (=Ravin of Dumortier 1871) with siliceous sponges and crinoids. The locality of La Clapouze near St-Étienne-de-Boulogne contains a fauna similar to that of La Pouza. Roman (1950) placed the La Pouza and La Clapouze outcrops in the Bathonian, whereas Elmi (1967) and Alméras and Elmi (1996) placed it in the Lower Bathonian. The localities near La Voulte were recently discussed by Charbonnier et al. (2007) and Charbonnier (2009) who described the sponges and a number of crinoids from the Chénier Ravine, now placed in the Lower Callovian Gracilis Zone. This fauna is coeval with the famous La Voulte Lagerstätte (Ravin des Mines) containing a unique soft-bodied fauna, but largely devoid of crinoids (Etter 2002; Charbonnier 2009). The crinoids from La Pouza, La Clapouze and Chénier Ravine were described by Dumortier (1871) and de Loriol (1882), and those from the Chénier Ravine by Charbonnier et al. (2007) and Charbonnier (2009). The Bajocian/Bathonian fauna of Pont des Étoiles has never been discussed in detail. In the following overview, the original names of the authors cited are given.

Geological maps of the Ardèche margin along the SE Massif Central with fossil locations described in the text. Upper part, area between Aubenas and La Voulte with the locations of La Clapouze and La Voulte. Lower part, the four locations near La Voulte. Modified after Charbonnier (2009)
Pont des Étoiles
This outcrop is at a bridge of the D365 crossing the Lauvie brook and gorge. Crinoidal limestones of Late Bajocian/Early Bathonian age (Parkinsoni/Zigzag Zones) with mostly columnals, cirrals and some brachials of Isocrinus nicoleti are well exposed here. This type of rock is developed all along the vivaro-cevenole borderland. Alméras and Elmi (1996) assumed that the crinoid remains and the brachiopods were transported by currents or mass flows from adjoining platforms or swells and accumulated at the foot of the escarpment. Among the remains of I. nicoleti are additional taxa, including a new genus of Cyrtocrinida. The outcrop is also remarkable by the occurrence of three distinctive species of Balanocrinus.
La Pouza
West of Pont des Étoiles, the crinoidal limestones are surmounted by grey limestones and marlstone (“Calcaires gris de la Pouza”) of Early Bathonian age (Zigzag Zone, Alméras and Elmi 1996). Dumortier (1871) described the following species: Eugeniacrinus caryophyllatus Goldfuss, Eugeniacrinus fenestratus Dumortier, Eugeniacrinus nutans Goldfuss, Millericrinus sp. (two species), Pentacrinus pentagonalis Goldfuss and Pentacrinus subteres Goldfuss. In addition, Roman and Sayn (1928) mentioned Balanocrinus dumortieri de Loriol, Balanocrinus pacomei de Loriol, Cyclocrinus macrocephalus (Quenstedt), Eugeniacrinus aberrans de Loriol and Eugeniacrinus dumortieri de Loriol. De Loriol (1882–1889) described or mentioned Balanocrinus dumortieri de Loriol, Balanocrinus pacomei de Loriol, Balanocrinus subteres (Münster), Cyclocrinus macrocephalus (Quenstedt) (=one of Dumortier’s Millericrinus sp.), Eugeniacrinus aberrans de Loriol, Eugeniacrinus dumortieri de Loriol, Eugeniacrinus nutans Goldfuss, Pentacrinus dumortieri Oppel, Pentacrinus nicoleti Desor and Phyllocrinus fenestratus Dumortier.
A new track near the farm cutting through the entire Bathonian succession furnished material collected by A. S. Gale, in part from the surface, but mostly picked by the author from residues of approximately 100 kg of bulk material from three beds (“La Pouza 1-3”) full of echinoderm debris. A complete profile for the section was measured by A. S. Gale (Fig. 2), and he provided the following text (A. S. Gale, written communication, 2011): “The section exposed in trackside cuttings east of the farm buildings at La Pouza comprises about 17 m of thin limestones and marly clays dipping approximately south at 40°. The contact with the basement quartzites is not exposed and may be faulted. Two gaps in the succession, the lower one of just over 3 m and the upper of 2 m, are present. The lowest 5 m exposed comprise poorly fossiliferous marls, with scattered echinoid fragments and crinoid ossicles, alternating with 10–30 cm thick micritic limestones. The upper succession includes 15 thin limestones (mudstones–wackestones), one of which is the Limacean Marker Bed, because it includes numerous bivalves and echinoid debris. Four of the clay beds contain abundant debris of calcitic fossils, mostly echinoderms, and less frequently fragments of sponges (“La Pouza 0-3”). A number of the echinoderm fragments are abraded, suggesting accumulation in a high-energy environment. The debris is concentrated in the basal part of the clay beds, and becomes progressively sparser into the higher part of each bed. The alternating marl-limestone couplets appear to represent background sedimentation in relatively deep water, perhaps reflecting climatically driven cycles, with a low background input of calcitic detritus from shallower environments, probably washed downslope by storms. The four graded beds containing abundant echinoderm material represent substantial, brief, input from a shallower environment, perhaps generated by major storm events or slope instability. The consistent occurrence of these beds immediately overlying limestones, suggests that there is a relationship between processes controlling basin sedimentation and the influx of bioclastic debris.”

Stratigraphic section excavated by A. S. Gale during 2010 near the farmhouse of La Pouza. LaP 1–3 (Beds 1–3) were sampled and picked separately for the dominating species, Cyrtocrinus praenutans n. sp. (see Table 2)
Chénier Ravine (Ravin du Chénier)
Approximately, 20 m of marlstones and marly limestones are exposed in badlands 150 m SW of the famous La Voulte Lagerstätte (Charbonnier et al. 2007). A number of marly layers (details in Charbonnier 2009) furnished crinoids that are associated with sponges. The authors collected 16 crinoid cups, 13 Cyrtocrinus nutans (Goldfuss) and three Eugeniacrinites dumortieri (de Loriol). Including material from the collections of the University of Lyon, they listed 127 cups of the following species: Cyrtocrinus nutans (90 cups), Cyrtocrinus nutans voultensis (four cups), Gammarocrinites compressus (Goldfuss) (11 cups), Eugeniacrinites dumortieri and Phyllocrinus colloti de Loriol (9 cups each), Lonchocrinus sp. Jaekel and Dolichocrinus aberrans (de Loriol) (two cups each). Several hundreds of columnals and brachials were also mentioned, but not specifically assigned. Dumortier (1871) described only Eugeniacrinus caryophyllatus Goldfuss and Pentacrinus subteres Goldfuss. de Loriol (1882–1889) mentioned Pentacrinus dumortieri Oppel and Pentacrinus nicoleti Desor.
Mines Ravine (Ravin des Mines)
This is the La Voulte Lagerstätte, notable for its spectacular fauna of arthropods and soft-bodied cephalopods (Charbonnier 2009). Echinoderms are represented by the common ophiuroid Ophiopinna elegans (Heller) (see Hess 1960) and the rare comatulid Rhodanometra lorioli Manni, Nicosia and Riou (1985). In addition, Manni et al. (1985) figured an undetermined isocrinid pluricolumnal and remains of a second species of comatulid, also undetermined. Charbonnier et al. (2007) and Charbonnier (2009) explained the striking difference between the Chénier Ravine and Mines Ravine faunas by the autochthonous nature of the latter. In contrast, at least part of the Chénier Ravine fossils, such as the cyrtocrinids, are thought to have been transported from higher parts of the slope. Whereas Ophiopinna elegans certainly was autochthonous, the rare comatulids and the isocrinid may have been parautochthonous.
La Clapouze
Dumortier (1871) described the following species: Eugeniacrinus caryophyllatus Goldfuss, Eugeniacrinus fenestratus Dumortier, Eugeniacrinus nutans Goldfuss, Millericrinus sp., Pentacrinus cingulatus Münster and Pentacrinus subteres Goldfuss. de Loriol (1882) described or mentioned Balanocrinus dumortieri de Loriol, Balanocrinus subteres (Münster), Cyclocrinus macrocephalus (Quenstedt), Eugeniacrinus aberrans de Loriol, Eugeniacrinites dumortieri de Loriol, Eugeniacrinus nutans Goldfuss, Cyclocrinus macrocephalus (Quenstedt), Phyllocrinus fenestratus Dumortier and Pentacrinus dumortieri Oppel. Charbonnier (2009, p. 160) mentioned the following species: Eugeniacrinus caryophyllites, Cyrtocrinus nutans, Phyllocrinus fenestratus, Dolichocrinus aberrans, Balanocrinus subteres and Pentacrinus cingulatus.
Materials and methods
The material collected by the author in the late 1950s includes specimens from the four sites mentioned above. In the following (Table 1), revised names are given. For details, see section “Systematic palaeontology”.
Repository, Natural History Museum Basel (Switzerland).
The La Clapouze location, described in some detail by Roman (1950, p. 47), is at present not well exposed and is impoverished in crinoids (A. S. Gale, personal communication, 2011).
The material collected in bulk at La Pouza by A. S. Gale was picked by the author for the crinoids, ophiuroids and asteroids; the asteroids are to be described by A. S. Gale in a separate paper and the relatively rare ophiuroids are studied by Ben Thuy. There are no essential differences in the crinoid material from the three beds sampled, except for the rare comatulids that were found only in one or the other bed.
Taphonomy of the La Pouza site
Fossil content of the three claystone beds with abundant debris of echinoderms is essentially identical. Crinoids are by far the most abundant fossils with nearly 100,000 ossicles, dominated by remains of the cyrtocrinid Cyrtocrinus praenutans n. sp. Preservation is mostly good to excellent, and the number of abraded or corroded ossicles, mainly cups of Cyrtocrinus praenutans n. sp., is rather small. With the exception of pinnulars of Cyrtocrinus praenutans n. sp., all types of ossicles and all sizes are present, down to postlarval columnals and brachials of less than 1 mm. Pinnulars may have been present in the finest fraction that was picked only cursorily. In order to examine if certain types of ossicles are preferentially preserved, those of the dominant species Cyrtocrinus praenutans n. sp. were counted in each bed (Table 2).
The results demonstrate that there are no fundamental differences in deposition and preservation of ossicle types between the three beds, although numbers do not correspond exactly. The large percentage of single radials, radial pairs and radial triplets (see Appendix 1) is unusual, because cyrtocrinid cups are connected by tight synostosis with interlocking grooves (Fig. 14f–g). Indeed, single radials exceed intact cups in Bed 1 (see Appendix 1). In contrast, isolated radials of the similar species Cyrtocrinus nutans (Goldfuss) are exceptional in Middle Oxfordian outcrops of Germany and Switzerland. Despite the richness of the material at La Pouza, only a single pluricolumnal of Cyrtocrinus praenutans n. sp. was found and only a single case of a first secundibrachial still attached to the second primibrachial is noted (Fig. 16e). The relatively smaller percentage of attachment discs or holdfasts may be explained by their fixation to the bottom. Surprisingly, first primibrachials number only approximately a third of second primibrachials. This is unexpected because both ossicle types are easily recognized, and their number should correspond. First primibrachials are lower and blade-like in comparison with the mostly sturdy and angular second primibrachials. Thus, they may have been more susceptible to winnowing. However, such a sorting process would also have included the ossicles from the fine fraction (see Appendix 2) where first and second primibrachials are equally represented.
The relatively small number of brachials and the lack of pinnulars suggest some winnowing. I conclude from this that the animals lived on a hardground bottom, remained exposed for some time after death and were carried away before being embedded in the clay sediment where conditions for epizoans were unfavourable. This is indicated by the relatively few cases of ossicles with epizoans, such as serpulids (Fig. 24g). Such a scenario is supported by the small number of pluricolumnals of the isocrinids Isocrinus dumortieri and Balanocrinus dumortieri (ratios of pluricolumnals, mostly of two columnals to single columnals: 8 % in I. dumortieri and 4.5 % in B. dumortieri). In contrast, a ratio of 10.5 % has been observed in Balanocrinus ticinensis Hess from the Pliensbachian of Arzo (Hess 2006), deposited under similar conditions. However, the crinoid-rich marlstones at Arzo are underlain by the hardground thought to be the attachment site for the crinoids. This would explain the lesser degree of disarticulation. In contrast, the La Pouza clay beds were the burial ground of ossicles carried away from the living site.
Specimens reported in the literature (Dumortier 1871, de Loriol 1882–1889) and hand collected by the author in the late 1950s are from sedimentary rocks exposed near the La Pouza farmhouse. These marly limestones (“marno-calcaires gris, clairs” in Sayn & Roman 1928, p. 17; “couches marneuses blanchâtres, dures, très rugueuses” in Dumortier 1871, p. 5) are rich in crinoids, sponges and brachiopods; echinoids and asteroids are also common (“faune de la Pouza”, Sayn & Roman 1928, p. 31). The lithology is similar to that of the La Clapouze locality (“marnes durcies, grises, jaunâtres et claires”, Dumortier 1871, p. 6), and this is also true of the type of fossils, even though other crinoid species are dominant at the different localities. The three claystone beds sampled by A. S. Gale lack sponges and brachiopods, and echinoids and asteroids are rare in comparison with the dominant crinoids. The sedimentary rocks of the “faune de la Pouza”, with sponges and brachiopods, are similar to the Middle Oxfordian Birmenstorf Member of northern Switzerland and the somewhat younger Lochen Beds of the Swabian Alb (Hess & Spichiger 2001). Sediments of the Birmenstorf Member exposed in the Chalch quarry (Hess & Spichiger 2001) were assumed to have been deposited on a swell in moderately deep water, and those of the Lochen Beds deposited at the margins of sponge–algal bioherms (Ziegler 1977). Obviously, the large majority of the animals constituting the”faune de la Pouza” lived on hardgrounds similar to those of the Birmenstorf Member and are representative of the original fauna. Other components were likely to have been soft substrate dwellers. These include asteroids such as Tylasteria, and rare Terminaster and pterasterids (A. S. Gale, personal communication, 2012). Transport to the depositional area appears to have been limited. In contrast, the fossils isolated from the claystone beds are predominantly crinoids dominated by Cyrtocrinus praenutans n. sp., which certainly lived on hardground and not on soft bottom. The huge dominance of crinoids, the absence of sponges and brachiopods and also the low percentage of echinoids and asteroids may be due to preferred transport of the generally smaller crinoid elements and low density of their skeleton.
Systematic palaeontology
Remark. Taxonomy and terminology, including references to authors, after Hess and Messing (2011)
-
Order Isocrinida Sieverts-Doreck, 1952
-
Suborder Pentacrinitina Gray, 1842
-
Family Pentacrinitidae Gray, 1842
-
Pentacrinites Blumenbach, 1804
-
Pentacrinites ausichi n. sp., Fig. 3
Fig. 3 
Pentacrinites ausichi n. sp., Upper Bajocian–Lower Bathonian, Pont des Étoiles. a Nodal, holotype; a1, upper (proximal) facet; a2, lateral with cirrus socket, M10921. b Internodal, b1 upper facet, b2 lateral (note sculpturing), M10922. c Large internodal with smooth latus, M 10920. d1–2 Facets of oval cirral, M10981. e Facet of circular cirral, M10923. f Proximal cryptosyzygial facet of epizygal brachial, M10924. Scale bars 1 mm

Material. 6 columnals (one nodal), 5 cirrals, 3 brachials from Pont des Étoiles.
Holotype. Nodal, Fig. 3a, M10921.
Paratype. Internodal, Fig. 3b, M10922.
Etymology. In honour of William I. Ausich, foremost authority on fossil crinoids.
Type locality and horizon. Pont des Étoiles, La Voulte-sur-Rhône (Ardèche, France), Late Bajocian/Early Bathonian Parkinsoni/Zigzag Zones.
Diagnosis. Nodals and internodals of similar outline, but nodals higher than internodals; cirrus sockets centrally placed on latus, not displaced interradially; symplectial areolae narrow, elevated, straight-sided to elliptical.
Description. The columnals are pentagonal, with slightly concave latus. The only known nodal is slightly weathered, robust, rather high and has two similar facets with elevated, nearly straight-sided areolae. The large cirrus sockets occupy most of the latus; they are centrally placed and the cirral scar has a low transverse ridge slightly thickened at the ends. The large internodal (Fig. 3c) is low (height 2.2 mm), the areolae are straight-sided and have distinct crenulae on the elevated margins, the radial areas are nearly smooth but have a few tubercles towards the margin, and the latera are smooth. The smaller internodal (Fig. 3b) has elliptical areolae and radial areas with tubercles towards the latera that are ornamented by tubercles. In both internodals, the small lumen is surrounded by a small elevated perilumen. The three internodals not figured have diameter/height ratios of 9.5/2.6, 8.0/2.0 and 6.0/2.4 mm. The five cirrals are circular to slightly oval, and the ligament pits are nearly equal (Fig. 3e). One cirral ossicle (Fig. 3d) has one facet with a transverse ridge that matches a corresponding depression on the other facet. Similar cirrals are known from the proximal part of cirri of Pentacrinites dargniesi. A nearly circular, thin epizygal brachial has a proximal cryptosyzygial facet (Fig. 3f), while the other side is muscular with a pinnule socket. The cryptosyzygial facet resembles facets of P. dargniesi, supporting the assignment to a species of Pentacrinites. The two other brachials are similar in outline, but both facets are muscular, the distal one with pinnule socket.
Remarks. This species differs from the coeval Pentacrinites dargniesi Terquem & Jourdy by the presence of well-developed internodals whose size is similar to that of the corresponding nodal. Small internodals were not detected, but pluricolumnals would have to be available to verify this. De Loriol (1888) described three additional Bajocian species of Pentacrinites. Two of them, Extracrinus (=Pentacrinites) lorteti and Extracrinus (=Pentacrinites) sorlinensis, are from the Bajocian of Saint-Sorlin (Ain); they differ from P. dargniesi mainly by smooth brachials and cylindrical cirrals; the two species may well be conspecific. The columns from Saint-Sorlin are densely cirrated and, thus, seem to be devoid of larger internodals. The third species, Extracrinus (=Pentacrinites) babeaui, occurs at numerous locations in France, but is known only from columnals; the large nodals and the small internodals are quite similar to those of P. dargniesi.
-
Suborder Isocrinina Sieverts-Doreck, 1952
-
Family Isocrinidae Gislén, 1924
-
Subfamily Isocrininae Roux, 1981
-
Isocrinus von Meyer in Agassiz, 1836
-
Isocrinus nicoleti (Thurmann in Thurmann & Étallon, 1861), Fig. 4
Fig. 4 
Isocrinus nicoleti (Thurmann in Thurmann & Étallon), Upper Bajocian–Lower Bathonian, Pont des Étoiles. a Pluricolumnal with complete noditaxis of eight columnals, a1 cryptosymplectial facet of infranodal on top, a2 lateral view, M10927. b Pluricolumnal with incomplete noditaxis, b1 symplectial facet, b2 lateral view, M10928. c Proximal (upper) facet of proximal nodal, M10929. d Proximal (upper) facet of nodal from mesistele, M10930. Scale bar 1 mm

1845 Pentacrinus Nicoleti, Desor, p. 5 (without description).
1861 Pentacrinus Nicoleti Desor; Thurmann & Étallon, p. 351 (first short description).
1879 Pentacrinus Nicoleti Desor; de Loriol, p. 139, pl. 15, figs. 34–36.
1887 Pentacrinus Nicoleti Desor; de Loriol, p. 165, pl. 154–161.
1972 Isocrinus nicoleti (Thurmann); Hess, p. 65, pl. 21, figs. 1, 2, 4.
1975 Isocrinus nicoleti (Thurmann); Hess, p. 55, pl. 7, fig. 4; pl. 11, 16.
Material. The bulk of the material consists of pluricolumnals from Pont des Étoiles where the species forms a crinoidal limestone. At La Pouza, the species is rare, and the columnals are smaller: one pluricolumnal of 6, one pluricolumnal of 5, one pluricolumnal of 4 (with nodal), one pluricolumnal of 3 (with infranodal), 7 pluricolumnals of 2 (2 with infranodal, 1 with nodal), 3 internodals, 2 nodals and 2 infranodals.
Diagnosis (column). Columnals are stellate to angular pentagonal, low, nodals slightly higher than internodals, cirrus sockets large, as high as nodal; latera of internodals are somewhat inflated between the margins, and inflated columnals may alternate with hardly inflated ones; noditaxis of 8 columnals.
Description. The material from Pont des Étoiles is largely composed of partly abraded pluricolumnals, but a number of well-preserved specimens are also available. They include complete noditaxes (Fig. 4a), incomplete noditaxes (Fig. 4b) and single columnals. The stellate columnals are low, and they alternate slightly in height and diameter. The latera are somewhat inflated between the margins; inflated columnals may alternate with hardly inflated ones (Fig. 4a2). The interradii are acute in smaller individuals (Fig. 4b), but become more rounded in larger specimens (Fig. 4a). The nodals are somewhat higher than the internodals, with cirrus sockets of equal height (Fig. 4a). Proximal nodals (Fig. 4c) are more strongly stellate than more distal nodals (Fig. 4d). The infranodals (top columnal of Fig. 4a) are slightly lower than the following internodals, and the proximal facet is cryptosymplectial (Fig. 4a1). All other columnal articulations are symplectial. The arrangement of the crenulae is typical of the genus, with similar radial and interradial crenulae.
Remarks. The figured specimens are from the Pont des Étoiles site; the few columnals from La Pouza have the same characters, but are smaller.
-
Isocrinus dumortieri (de Loriol 1877), Fig. 5
Fig. 5 
Isocrinus dumortieri de Loriol, Lower Bathonian, La Pouza. a Pluricolumnal of three elements, a1 upper facet, a2 lateral with continuous crest on upper columnal, M10900. b Facet of internodal, collar with strong lateral extensions, M10935. c Facet of internodal with jagged collar, M10987. d Facet of higher small internodal with collar broken into knobs, M10957. e Proximal (upper) facet of small nodal, M10933. f Facet of small low internodal with partly interrupted collar, M 10934. g Facet of juvenile, high internodal, M10938. h Proximal (upper) facet of nodal, M 10937. i Infranodal, i1 proximal (upper) facet, i2 distal (lower) facet, M10984. j Large internodal with continuous collar, M10936. Scale bars 1 mm

1865 Pentacrinus dumortieri Oppel, p. 317
1887 Pentacrinus dumortieri Oppel; de Loriol, p. 174, 182, pl. 162, figs. 1–8
Material. Pont des Étoiles. Pluricolumnals: one of 11 (with nodal and infranodal = noditaxis), one of 9 (with nodal and infranodal = noditaxis), one of 8 (with nodal and infranodal = noditaxis), two of 7 (one with nodal and with infranodal = noditaxis), one of 6 (with infranodal). La Pouza. Pluricolumnals: one of 8 (with nodal), two of 5 (one with infranodal), five of 3 (one with nodal), 14 of 2 (8 with nodal and 3 with infranodal, smallest has a diameter of 1 mm); 29 nodals, 73 infranodals, 173 internodals (smallest diameter = 0.8 mm), 1 topmost columnal.
Diagnosis. Columnals mostly low, stellate to pentagonal or pentalobate, with slightly concave latus, or circular; articular facets of large internodals with elliptical petals and adradial crenulae that may converge towards the lumen; adradial crenulae of smaller, circular internodals reduced to bands towards lumen; nodals with large cirrus sockets, not sunken, directed slightly upwards and surrounded by rim; width of cirral scar 50 % of nodal diameter, cirral scar with tubercle at each end; latera of internodals with median ridge that may be jagged or interrupted into segments or reduced to tubercles or knobs.
Description. The material from La Pouza includes columnals of all sizes and excellent preservation (Fig. 5). Among them are small, more or less circular nodals and internodals with adradial crenulae reduced to V-shaped element (Fig. 5d, f, g). The median ridge on the latera may be sharp and continuous (Fig. 5c, i, j), or interrupted with prominent processes (Fig. 5b), or the ridge may be modified into granules or knobs (Fig. 5a, d–h). The pluricolumnal figured combines these features (Fig. 5a2); the top columnal has a continuous ridge, the middle one a ridge interrupted and the lower one a band of tubercles.
Remarks. The species name, dumortieri, was by Oppel (1865) and without description. The species was first characterized by de Loriol (1877), who mentioned noditaxes varying between 9 and 13. In the material from Pont des Étoiles, the noditaxis varies between 7 and 11. The material from La Pouza is more disarticulated and has not furnished a pluricolumnnal consisting of a noditaxis. Isocrinus dumortieri is easily distinguished from I. nicoleti by the prominent median ridge on the latus of most columnals, including juveniles.
-
Subfamily Balanocrininae Roux, 1981
-
Balanocrinus Agassiz in Desor, 1845
-
Balanocrinus dumortieri de Loriol 1877, Fig. 6
Fig. 6 
Balanocrinus dumortieri de Loriol, Lower Bathonian, La Pouza. a Pluricolumnal of four columnals, a1 lateral with nodal, a2 upper facet of internodal, M10843. b Facet of internodal with marginal rim, M10932. c Facet of internodal with well-developed adradial crenulae, M10982. d Facet of pentagonal, low proximal internodal with ribbon-like adradial crenulae, M10988. e Juvenile pluricolumnal, e1 facet, e2 lateral, M10986. f Internodal with pronounced rim, f1 facet, f2 lateral, M10983. g Distal (lower) facet of tetramerous nodal, note stereomic overgrowth and epizoic foraminifer, M 10989. h Facet of tetramerous internodal, M10917. Scale bar 1 mm

1871 Pentacrinus pentagonalis (Goldfuss); Dumortier, p. 46, pl. 5, figs. 1–3.
1887 Balanocrinus dumortieri de Loriol, p. 324, pl. 187, figs. 3–8.
1996 Balanocrinus dumortieri de Loriol; Klikushin, p. 117, pl. 8, figs. 1–2.
2007 Balanocrinus dumortieri de Loriol; Charbonnier et al., figs. 10e–f.
Material. Pont des Étoiles: one pluricolumnal of 3. La Clapouze: 4 internodals. Chénier Ravine: 3 internodals. La Pouza (collected by the author): 8 pluricolumnals: one of 8 (with nodal in the middle), two of 6 (one with infranodal), one of 4, two of 3 (one with infranodal), two of 2 (one with nodal) and 10 internodals. La Pouza (Beds 1–3): 2326 internodals, 231 nodals (the smallest with a diameter of 0.7 mm), 259 infranodals; and the following pluricolumnals: one of 6 (with nodal), two of 5 (one with nodal), 9 of 4 (4 with nodal), 15 of 3 (2 with infranodal), 100 of 2 (15 with nodal, 7 with infranodal).
Diagnosis. Facets of internodal columnals circular to subcircular to weakly pentalobate or rounded pentagonal, with rather long marginal crenulae; crenulae commonly V-shaped adradially, where size of crenulae decreases towards the lumen, or crenulae are modified into elevated bands; latera concave and smooth in circular columnals, while in pentalobate columnals commonly a tubercle in each radius; latus separated from facet by sharp ridge; nodals somewhat higher than internodals, with large cirrus sockets nearly as high as nodal, separated by tubercle.
Description. The columnals are mostly circular, tending to slighly pentalobate or rounded pentagonal. The facets of internodals have rather pronounced marginal crenulae that are commonly V-shaped adradially (Fig. 6a1, b). The radial bands of crenulae may be flat (Fig. 6a1, b) or elevated (Fig. 6d, e1). The distinct marginal ridges are thickened in rare cases (Fig. 6f). Distal facets of nodals and proximal facets of infranodals are cryptosymplectial; Fig. 6g shows the lower (distal) facet of a tetramerous nodal with stereomic growth sealing the axial canal, possibly at the end of the column. Tetramerous columnals occur occasionally (Fig. 6h). Juvenile columns are essentially similar to adult ones (Fig. 6e). The smallest nodal has a diameter of 0.8 mm, and the smallest internodal of 0.5 mm. The latera of circular columnals are smooth, but those of pentalobate outline commonly have a rounded tubercle at each radius. This is the commonest isocrinid at La Pouza, by far represented by numerous pluricolumnals and columnals. No intact noditaxis has been found by the author, but can be estimated from the number of nodals or infranodals and internodals at approximately 11 columnals. De Loriol (1887, p. 325) mentions a noditaxis of 9 columnals.
Remarks. The species is easily recognized by the concave latera of the internodals, separated from the facets by a sharp ridge. Brachials assigned to this species include distinctive syzygies (Fig. 9b–d). Syzygies are not uncommon in secundibrachials of Balanocrinus species (Hess 2006, pl. 25, figs. 10, 12–14). The present brachials also resemble brachials from the Albian of England (Hess & Gale 2010, fig. 6x) that were unassigned, but may have belonged to Balanocrinus smithi Hess.
-
Balanocrinus pacomei de Loriol 1877, Fig. 7d–g
Fig. 7 
Balanocrininae from Upper Bajocian–Lower Bathonian, Pont des Étoiles (a–d, f–g) and Lower Bathonian, La Pouza (e). a–c Balanocrinus inornatus (d’Orbigny); a pluricolumnal of three elements, a1 upper facet, a2 lateral, M10916; b pluricolumnal of two, b1 upper facet, b2 lateral with cirrus sockets, M10914; c juvenile pluricolumnal of two, c1 upper facet, c2 lateral, M10915. d-g Balanocrinus pacomei de Loriol; d pluricolumnal of 15 internodals, d1 upper facet (infranodal, note tetramerous symmetry), d2 lateral, d3 lower facet, M10911; e small internodal, e1 facet, e2 lateral, MM10985; f pluricolumnal of two internodals, f1 upper facet, f2 lateral, M10913; g nodal, g1 lateral, g2 proximal (upper) facet, M10912. Scale bars 1 mm

1887 Balanocrinus Pacomei de Loriol, p. 318, pl. 186, figs. 1–6.
Material. Pont des Étoiles. 22 pluricolumnals: one of 15 (without nodal or infranodal), two of 8, two of 7, four of 6 (one with nodal), two of 5, three of 4, four of 3, four of 2 (one with nodal); one nodal and 11 internodals. At La Pouza, the species is rare: one pluricolumnal of 2 (without nodal or infranodal), a single internodal and a single nodal.
Diagnosis. Columnals circular, with slightly convex, smooth latera, facets with short marginal crenulae and narrow elevated granular radial bands; nodals with large cirrus socket nearly as high as nodal, cirral scar with narrow transverse ridge interrupted by pore.
Description. The columnals are all similar. A pluricolumnal of 9 has a nodal distally, and the facet of the proximalmost internodal is symplectial. This indicates a noditaxis of more than 10 columnals, which is confirmed by a pluricolumnal of 15 internodals with an infranodal on top (Fig. 7d). Tetramerous columnals occur occasionally (Fig. 7d).
Remarks. The species is easily distinguished from the other species of the genus by the more or less convex latera and the adradial crenulae developed into elevated bands.
1850 Pentacrinus inornatus d’Orbigny, p. 891.
1887 Balanocrinus inornatus d’Orbigny, de Loriol, p.311, pl. 184, figs. 3–9.
1911 Balanocrinus inornatus d’Orbigny, Boule, p. 111 (pl. 22, fig. 12, published 1911).
1912 Balanocrinus inornatus d’Orbigny, Lissajous, p. 173, pl. 18, fig. 32
1927 Balanocrinus inornatuss (d’Orbigny), Valette, p. 27.
Material. Pont des Étoiles: one pluricolumnal of 2 with nodal (Fig. 7b), one pluricolumnal of 3 (Fig. 7a), four pluricolumnals of 2 (Fig. 7c). La Pouza: one internodal. Chénier Ravine: three internodals.
Diagnosis. Large species, columnals high, cylindrical, with smooth latus; facets with short marginal crenulae and weak radial ridges, so that internodal articular surface may resemble cryptosymplexy; cirrus sockets hardly sunken, half as high as nodal, width of cirral scar approximately 40 % of nodal diameter, transverse ridge modified into two isolated weak tubercles.
Description. The columnals are all similar. The ratio of height to diameter varies from approximately half in adult specimens to one in the juvenile specimen (Fig. 7c).
Remarks. The species was first mentioned by d’Orbigny (1850), and described and figured in some detail by de Loriol (1877) on the basis of material from the Oolithe blanche (now considered lower Bathonian) of Normandy, Creuse and Orne. D’Orbigny’s type, a pluricolumnal of 6 without a nodal, is from Guéret (Sarthe), and it was figured by Boule (1911) in the Types du Prodrome. According to de Loriol (1877), the column seems to be pentagonal near the cup; more distally, it is cylindrical with circular facets. The species was mentioned or described from several locations of France and Spain. Lissajous (Lissajous 1912, pl. 18, fig. 32) figured a small pluricolumnal from the Bajocian Parkinsoni Zone of Mâcon. The species was described, but not figured, by Valette (1927) from the upper Bajocian of Catalogne (Tivenys). Balanocrinus inornatus is easily distinguished from the two other species of Balanocrinus occurring at Pont des Étoiles by the smooth, compact nature of the cylindrical columnals. Valette mentioned that the articular facets are commonly indistinct (“la facette articulaire de cette espèce est assez souvent peu accentuée, et semble parfois disparaître”). Smooth circular columnals are distinctive for B. subteres (Münster in Goldfuss), the type species of the genus from the Oxfordian, but the radial ridges are more prominent and have larger crenulae; the cirral scars are more pronounced.
Remains of isocrinids that cannot be assigned to a given species
Isocrinid cirrals
La Pouza (1-3): 136, 1 pluricirral of four. The cirrals are elliptical to nearly circular in section and vary from short to long, with concave body. Facets are similar and synarthrial. The cirrals are mostly smooth, but short, circular cirrals have fringed borders. Assignment to a given species is not possible, although the ornamented ossicles may have belonged to Isocrinus dumortieri.
Isocrinid brachials (Figs. 8, 9)
The material isolated from the from the La Pouza samples yielded a considerable number of isocrinid cup plates and brachials with distinctive characters. Among them are a few radials and primibrachials, but mainly secundibrachials with different types of articulation facets. Two major groups may be distinguished.

Radial (a) and brachials (b–k) of Isocrinida, Lower Bathonian, La Pouza. a Radial, a1 proximal (lower) with facets to basals, a2 distal, M10853. b Secundibrachial with symmorphy, b1 adoral, b2 distal (cryptosyzygial to synostosial facet), M 10856. c Distal facet of first primibrachial with verrucose aboral surface, M10888. d Proximal facet of first primibrachial, M 10889. e Cryptosyzygial facet of secundibrachial with verrucose aboral surface, M10882. f Distal muscular facet of secundibrachial with verrucose aboral surface and pinnule socket, M10890; g Axillary second primibrachial, g1 proximal facet (cryptosyzygy), g2 distal (muscular facets), M10948. h Distal muscular facet of secundibrachial with pinnule socket, M10886. i Distal muscular facet of wide secundibrachial with pinnule socket, M10946. j Epizygal secundibrachial, j1 proximal facet with syzygy restricted to margin, j2 distal muscular facet with pinnule socket, M10949. k Wide secundibrachial with flat relief, M10945. Scale bars 1 mm

Secundibrachials with syzygies and cryptosyzygies of Isocrinida, Lower Bathonian, La Pouza. a Cryptosyzygy, M10878. b Proximal facet of epizygal (other facet is muscular with pinnule socket), M10885. c Epizygal, c1 proximal facet, c2 distal facet with pinnule socket (arrow), M10991. d Epizygal, d1 proximal facet, d2 distal facet with pinnule socket (arrow), M10883. e Cryptosyzygy (other facet is muscular without pinnule socket), M 10892. Scale bars 1 mm
Ossicles with smooth latus, and brachials more or less circular in outline, except elongate distal secundibrachuials which are elliptical
Most of the ossicles may be tentatively assigned to Balanocrinus dumortieri de Loriol. Not only is this species the prominent isocrinid at La Pouza, but the shape of the secundibrachials with muscular facets resembles those of other species of the genus (Hess, research in progress). In addition, symmorphies and especially syzygies also occur in Balanocrinus.
Radials: 12 (Fig. 8a), the facet to the basals (Fig. 8a1) indicates that the basals were in contact, a character typical of Balanocrinus. The aboral surface is smooth in most cases, but verrucose in two radials.
First primibrachials: 35. The distal facet is synarthrial or cryptosynarthrial.
Axillaries: Sixty-one rather high axillaries with a proximal synostosial or symmorphial facet appear to be second primibrachials (Fig. 8g); two low axillaries with proximal synarthry certainly are second primibrachials, matching the first primibrachials in Fig. 8c; 118 axillaries with muscular proximal facet are secondary axillaries (secundibrachials). True isocrinids have primibrachials articulated by synarthries as in Fig. 8c. The rather numerous high axillaries with smooth ligamentary proximal facet (Fig. 8g) cannot be classified with certainty at present.
Secundibrachials. (1) The large majority of secundibrachials, 3,872 in number, have two muscular facets, the distal with pinnule socket (Fig. 8h). Most of these ossicles are thought to belong to Balanocrinus dumortieri. (2) Secundibrachials with syzygy: 149, 49 of which are hypozygal (proximal facet muscular, distal facet syzygial) and 100 are epizygal (proximal facet syzygial, distal facet muscular with pinnule socket (Fig. 9b–d). Syzygial brachials generally are low. Syzygies are common in comatulids, but remains of those are so rare that such assignment can be ruled out. Syzygies are not known from Isocrinus, but may occur in Balanocrinus, the common isocrinid at La Pouza. (3) Secundibrachials with cryptosyzgy or synostosis developed as symmorphy: hyposymmorphial (proximal musculer facet, distal cryptosyzygy), 82; episymmorphial (proximal cryptosyzygial, distal muscular with pinnule socket), 105 (Fig. 8b). Symmorphies (Fig. 8b) are characteristic of Isocrinus, and weak symmorphies or cryptosyzygies (Fig. 9e) are found in Balanocrinus. (4) Mostly rather high secundibrachials, with flat cryptosyzygy or synostosis that may approach symmorphy (the distinction is not sharp in some cases): muscular facet (proximal) without pinnule socket, 116 (Fig. 9e); muscular facet (distal) with pinnule socket, 190. Ossicles of this group may be assigned to Balanocrinus. (5) Secundibrachials with both facets synostosial or cryptosyzygial (Fig. 9f): 2. Ossicles in group 5 seem to belong to the same species as those in group 4.
Pinnulars: 27. The pinnulars are of different length, laterally compressed and triangular in section. They cannot be assigned to a specific isocrinid.
Ossicles with verrucose or granular latus
Secundibrachials belonging here are mostly verrucose or have a jagged edge, but some are also smooth. They are generally low and wide, while some are more or less rectangular (Fig. 8f), orally prolonged (Fig. 8e, j) or trapezoidal (Fig. 8i, k). Their assignment is doubtful, but they may belong to Isocrinus dumortieri de Loriol, a species characterized by ornamented columnals.
Radials: 2.
First primibrachials: 5 (Fig. 8c, d)
Second primibrachials: 6 (low)
Secundibrachials with proximal cryptosyzygy and distal muscular facet with pinnule socket: 11 (Fig. 8e); secundibrachials with proximal muscular facet and distal cryptosyzygy or syzygy without pinnule socket: 15; secundibrachials with two cryptosyzygial or synostosial facets: 1; secundibrachials with two muscular facets, the distal one with pinnule socket: 300 (Fig. 8f, i, k).
-
Order Comatulida A. H. Clark, 1908
-
Suborder Comatulidina A. H. Clark, 1908
-
?Superfamily Paracomatuloidea Hess, 1951
-
?Family Paracomatulidae Hess, 1951
-
Singillatimetra n. g.
Etymology. From singillatim (latin, single) and metra, the suffix commonly used for comatulids; the centrodorsal is composed of a single, disc-like element.
Diagnosis. Centrodorsal low, asymmetrical in outline; 5 bulging cirrus sockets arranged irregularly; no radial cavity or axial canal; aboral and adoral sides similar, weakly sculptured, on aboral side irregular tubercles and weak radial impressions without crenulae, on adoral side narrow interradial bands with short crenulae for articulation to basals.
Type species. Singillatimetra inordinata n. sp.
Remarks. This peculiar centrodorsal has properties of the Paracomatuloidea, but consists of only one piece. The lack of an axial canal indicates that it was not part of a centrodorsal composed of several pieces. The arrangement of the cirrus sockets, offset to either side of the radial midline, and the sign of crenulae on the adoral side suggest a paracomatuloid relationship. The form is tentatively assigned to this superfamily.
-
Singillatimetra inordinata n. sp., Fig. 10a
Fig. 10 
Centrodorsals of comatulids, Lower Bathonian, La Pouza. a Singillatimetra inordinata n. g., n. sp., holotype; a1 aboral, a2 lateral, a3 adoral, M10838. b Andymetra galei n. g., n. sp., holotype; b1 aboral, b2 lateral, b3 adoral, M10841. c Palaeocomaster cf. guirandi de Loriol, c1 aboral, c2 lateral, c3 adoral, M10837. Scale bars 1 mm

Material. Only the holotype is available.
Holotype. Centrodorsal, diameter 2.6 × 3 mm, height 0.9 mm, M10838.
Etymology. After the irregular arrangement of the cirrus sockets on the centrodorsal.
Type locality and horizon. Early Bathonian (Zigzag Zone), La Pouza, La Voulte-sur-Rhône (Ardèche, France).
Diagnosis. See genus (monotypic).
Description. The centrodorsal is asymmetrical in outline, with five bulging cirrus sockets offset from the radial midlines. The cirral scar is unsculptured, except for a weak transverse ridge in the upper part separating an upper third at an angle of 120° from a lower two-thirds. Both facets are similar and weakly sculptured, without radial cavity or axial canal. The presumed aboral side has some irregular impressions and tubercles, the presumed adoral side shows five rather regular narrow bands flanked by short crenulae or tubercles This structure is thought to have been articulated to basals.
-
Superfamily Solanocrinitoidea Jaekel, 1918
-
Family Solanocrinitidae Jaekel, 1918
-
Solanocrinites Goldfuss, 1829
-
Solanocrinites voultensis n. sp., Fig. 11c
Fig. 11 
Comatulids, Lower Bathonian, La Pouza. a-b Palaeocomaster messingi n. sp.; a centrodorsal, paratype, a1 aboral, a2 lateral, a3 adoral, M10840; b cup with centrodorsal, holotype, b1 aboral, b2 lateral, b3 distal (adoral), M10842. c Solanocrinites voultensis n. sp., centrodorsal, holotype; c1 aboral, c2 lateral, c3 adoral, M10841. Scale bars 1 mm

Material. Only the holotype is available.
Holotype. Centrodorsal, diameter 4.2 × 4.6 mm, height 2.4 mm, M10844.
Etymology. After the area of the type locality.
Type locality and horizon. Early Bathonian (Zigzag Zone), La Pouza, La Voulte-sur-Rhône (Ardèche, France).
Diagnosis. Centrodorsal columnar, aboral apex flattened, with narrow central depression and interradial ridges; cirrus sockets arranged in somewhat irregular 15 vertical columns of two to three rows, the five columns of each side separated by interradial ridges; narrow ridges for basal rods on the adoral side separated by weak irregular furrows, and the centrodorsal cavity a tenth of centrodorsal diameter.
Description. The centrodorsal is columnar and weakly pentagonal. Aboral and adoral sides are similar, but narrow bands for articulation to the basal rods and a moderately deep, narrow centrodorsal cavity (Fig. 11c3) leave no doubt about the nature of the upper facet. In contrast, the aboral side has five narrow interradial bands with slightly concave profile, three of the interradial sections are raised in folds with sockets at the end alternating with irregular furrows (Fig. 11c1). The five sides are covered by more or less circular cirrus sockets separated by interradial ridges. On each side, the sockets are arranged in five columns of mostly two to three rows; the sockets are displaced against each other, so that some protrude onto the aboral side, causing an irregular relief. The scars are unsculptured, but for a faint transverse ridge. The adoral side is dominated by the ridges for the narrow basal rods; between the ridges are shallow furrows, but also somewhat elevated areas, corresponding to slightly protruding cirrus sockets. The radial cavity is narrow.
Remarks. Assignment of the centrodorsal to Solanocrinites is unproblematic. In the similar genus Palaeocomaster, the cirrus sockets are arranged in irregular marginal circles without being separated by interradial ridges. This character also separates S. voultensis n. sp. from Palaeocomaster cf. guirandi (described below). Palaeocomaster schlumbergeri (de Loriol 1888, Bigot 1938) from the Bathonian of Calvados has a low, conical centrodorsal that may have variable interradial ridges (atypical of the genus), and there are only two columns of sockets in irregular rows of one to three. In addition, P. schlumbergeri has distinct impressions on the adoral side for the basals, which protrude on the outside and are united centrally, giving a star-like appearance. Palaeocomaster stellatus Gislén (1924) from the Bathonian of England is similar to P. schlumbergeri, but has a flat aboral side with a stellate impression.
Material. One centrodorsal. M10837.
Description. The centrodorsal is flattened, nearly circular, with a diameter of 4.0 mm and a height of 1.5 mm. The circular, prominent cirrus sockets are alternating and arranged in two irregular rows not interrupted by interradial ridges. The aboral side is somewhat weathered, cirrus free, irregular and has a central cavity of similar diameter (0.5 mm) as the centrodorsal cavity on the adoral side; smaller pits are due to weathering. The adoral side shows some flat interradial ridges for the basals.
Remarks. De Loriol (1888, p. 467; pl. 216, fig. 5) described a single centrodorsal from the Oxfordian of Le Pontet near Saint-Claude (France) under the name of Antedon guirandi. He also described a centrodorsal with cup from the same locality as Actinometra guirandi (de Loriol 1888, p. 535; pl. 227, fig. 2) without making reference to the earlier. Gislén (1924, p. 142) chose the latter specimen as type of guirandi, type species of Palaeocomaster Gislén, obviously considering the two specimens conspecific. The single centrodorsal has a diameter of 9 mm, whereas the centrodorsal with cup has one of only 3 mm. Both specimens have a low, flat centrodorsal, sculptured on the aboral side of Antedon guirandi, but smooth in Actinometra guirandi. Cirrus sockets of the latter are circular and surrounded by a rim, whereas they are more elliptical with a transverse ridge on the larger centrodorsal. Number and arrangement of sockets are similar in the two specimens. The present centrodorsal differs from that described by de Loriol (1889, pl. 216, fig. 5) by smooth cirrus sockets and a wide centrodorsal cavity on the adoral surface (narrow in de Loriol’s specimen). However, this difference and also different sculpturing may be due to weathering of the present specimen.
-
Palaeocomaster messingi n. sp., Fig. 11a, b
Material. Cup with centrodorsal (holotype), centrodorsal with attached basals (paratype).
Holotype. Cup with centrodorsal; largest diameter 5 mm, height 5 mm, height of centrodorsal 1.5 mm; M10842, Fig. 11b.
Paratype. Centrodorsal with basals; diameter 3 mm, height approximately 2 mm; M10840, Fig. 11a.
Etymology. In honour of Charles G. Messing for his work on living comatulids.
Type locality and horizon. Early Bathonian (Zigzag Zone), La Pouza, La Voulte-sur-Rhône (Ardèche, France).
Diagnosis. Centrodorsal low discoidal, irregular in shape, aboral apex smooth, interradial angles not produced; cirrus sockets crowded, in irregular on to four marginal cycles; five narrow basals hardly visible from outside, united centrally around narrow centrodorsal cavity; surface of radials not exposed, overhanging; radials with wide, steep facets, interarticular ligament fossae triangular and distinct, adoral muscle fossae high, separated by median notch; radial cavity moderately large.
Description. The centrodorsals of the two specimens vary in size; the centrodorsal of the holotype is reduced in diameter and height in comparison with the paratype. The centrodorsal is of irregular shape. Seen aborally, radial pairs of sockets with their base are fused and raised in folds, interrupted by more or less pronounced radial furrows, but this arrangement is somewhat irregular. The cirrus sockets vary in size and are unsculptured. The basals are narrow and united centrally, resembling the spokes of a wheel. They are visible from the outside as small knobs. The radial circlet is higher than the centrodorsal and does not display a free surface, which, thus, is overhanging. The steep radial articular facets are divided into three parts of roughly equal size, (1) an aboral ligament area with rather shallow fossa, (2) a middle part around the wide elliptical axial canal with transverse ridge below and lateral ridges containing triangular interarticular ligament fossae and (3) an upper part dominated by the muscle fossae separated by a deep notch. The radial cavity is moderately large.
Remarks. Palaeocomaster messingi n. sp. differs from P. schlumbergeri (de Loriol) by a much higher radial circlet with large axial canal and a centrodorsal of irregular shape with smooth aboral apex. In addition to the holotype of P. schlumbergeri from Calvados, numerous remains have become available from a site in the same department described by Bigot (1938). Bigot figured centrodorsals of more conical shape, with rather narrow aboral apex and cirrus sockets extending to near the apex, especially in smaller individuals. As in de Loriol’s holotype, the radial circlet is low compared with the present material, and the free surface of the radials is visible band-like above the centrodorsal. Topotype specimens in the Basel Museum, among them a well-preserved centrodorsal with cup and base of arms (M11034), display the same characters and are easily distinguished from the present specimen. The present material invites comparison with the Lower Jurassic (Hettangian) Palaeocomaster styriacus Kristan-Tollmann, the oldest comatulid with a centrodorsal composed of a single element. This species and the Pliensbachian P. morierei de Loriol (see Hess 2006) have centrodorsals composed of partly fused but distinct tubes that widen outwards to the cirrus socket. In P. messingi n. sp. such tubes are still visible, but fusion is more pronounced, and this is also true of the Upper Jurassic P. guirandi. Such morphology of the centrodorsal differs from that of Paracomatula species with rather compact centrodorsal, fused from a number of disc-like columnals. Singillatimetra inordinata n. g., n. sp., which may be a paracomatulid, also has a disc-like compact centrodorsal, without discrete “cirrus tubes”. Such differences suggest that morphology of centrodorsals in early comatulids is more complex than previously thought.
Andymetra n. g.
Diagnosis. Centrodorsal hemispherical, apex with only small cirrus-free area; cirrus sockets crowded, in several irregular rows, deep and hardly sculptured; adoral side with weak impressions of basals, centrodorsal cavity narrow, a tenth or less of centrodorsal diameter.
Etymology. In honour of Andy (Andrew) S. Gale for his contributions to the study of echinoderms; and -metra, suffix commonly used for comatulids.
Type species. Andymetra galei n. sp.
Remarks. The new genus shares the narrow centrodorsal cavity with the other genera of the family, but is distinct for its convex, hemispherical centrodorsal and the crowded cirrus sockets. These are arranged in irregular rows, not forming columns, as in Palaeocomaster, a genus with more or less flat aboral apex.
Other species. Antedon ladoixensis de Loriol (1888, p. 450; pl. 211, fig. 1) from the Bathonian of Côte-d’Or, France) and also reported from Calvados under the genus name Glenotremites (Bigot 1938).
-
Andymetra galei n. sp., Fig. 10b
Material. Only the holotype is available.
Holotype. Centrodorsal, diameter 3.7 mm, height 2 mm, M10841.
Etymology. Named after A. S. Gale who collected the material.
Type locality and horizon. Early Bathonian (Zigzag Zone), La Pouza, La Voulte-sur-Rhône (Ardèche, France).
Diagnosis. Centrodorsal nearly hemispherical, cirrus sockets crowded, in as many as four irregular rows.
Description. The centrodorsal is nearly hemispherical in profile and only the apex is devoid of cirrus sockets. These are crowded, deep and hardly sculptured; they occur in several, irregular rows, and columns cannot be distinguished. The adoral surface shows five weak flat ridges, some with faint crenulae; their shape and arrangement suggest that they served for reception of basal rods. The centrodorsal cavity is narrow.
Remarks. The present species has a more hemispherical centrodorsal than A. ladoixensis, which was assigned by Gislén (1924, p. 126) to the group of Glenotremites morierei (de Loriol); this Lower Jurassic species is now placed in Palaeocomaster (see Hess 2006, p. 59). The centrodorsal of A. ladoixensis (de Loriol 1888) is lower (diameter 4.5 mm, height 1.75 mm), with a more flattened apex. Bigot’s smaller centrodorsal (1938, pl. 4, fig. 21) is also low (diameter 2 mm, height 0.5 mm).
-
Order Millericrinida Sieverts-Doreck, 1952
-
Family Apiocrinitidae d’Orbigny, 1840
-
Apiocrinites sp., Fig. 23e
1871 Millericrinus; Dumortier, p. 46, pl. 5, figs. 4–6.
Material. La Pouza: two pluricolumnals of 4, one pluricolumnal of 2, 17 columnals (6 broken). La Clapouze: two pluricolumnals (one of 2 and one of 5 columnals).
Description. All columnals have straight latera and facets with rather strong radiating crenulae that commonly bifurcate near the margin (Fig. 23e). Most columnals have a slope between the crenularium and the lumen. The height/diameter ratio of columnals from La Pouza varies between 21 and 44 % in larger columnals, but reaches 70 % in small columnals of 1.3 and 3.8 mm in diameter. The lumen of larger columnals varies between 9 % and approximately 21 % of diameter; it reaches 40 % of the diameter in the smallest columnal (diameter 1.3 mm). The two pluricolumnals from La Clapouze with a diameter of 8.5 and 7.8 mm, respectively, are composed of low columnals with slightly convex latera; the lumen is relatively narrow at 10 % of the diameter.
Remarks. The columnals from the two sites are thought to belong to a single species despite the differences. Variable width of lumen is common in apiocrinitids. For example, width of the lumen in the Late Jurassic Apiocrinites roissyanus (d’Orbigny) from a single locality (Hess 1975, pl. 22, fig. 15–16) varies between 17 % (column diameter 9 mm) and 38 % (column diameters 5.3 and 8 mm). Assignment to Apiocrinites is tentative and is based on the occurrence of bifurcate and somewhat coarser crenulae. Such features are less common in otherwise similar columns of Liliocrinus Rollier (see Hess 1975, pl. 22, fig. 4–6). The large pluricolumnal (diameter 12 mm) figured by Dumortier (1871, fig. 5–6) from La Clapouze has a narrow lumen and strongly bifurcate crenulae (“Les lignes rayonnantes soint fortement dichotomes…”, Dumortier 1871, p. 47), and is similar to the present material.
-
Order Cyrtocrinida Sieverts-Doreck, 1952
-
Suborder Cyrtocrinina Sieverts-Doreck, 1952
-
Superfamily Eugeniacrinitoidea Zittel, 1879
-
Family Eugeniacrinitidae Roemer, 1855
-
Lonchocrinus Jaekel, 1907
-
Lonchocrinus dumortieri (de Loriol 1882), Fig. 12a–c
Fig. 12 
Cyrtocrinids, Lower Bathonian of La Clapouze (a–b, d–e) and La Pouza (c). a–c Lonchocrinus dumortieri de Loriol; a oblique lateral-distal view of cup, M10849; b adoral view of second primibrachial, M10850; c oblique adoral view of curved second primibrachial with blunted tip, M10994. d–e Phyllocrinus fenestratus Dumortier; d lateral view of adult cup with attached juvenile cup in oblique aboral view, M10863; e lateral view of cup with elongated radial on matrix, M10939. Scale bars 1 mm

1871 Eugeniacrinus caryophyllatus (Goldfuss); Dumortier, p. 48, pl. 5, fig. 12–13 (cup).
1882 Eugeniacrinus Dumortieri de Loriol, p. 132, pl. 14, figs. 1–12 (cups, second primibrachials, columnals).
1891 Eugeniacrinus Dumortieri (de Loriol); Jaekel 1891, p. 587, fig. 5 (second primibrachial); p. 647.
2007 Eugeniacrinus dumortieri (de Loriol); Charbonnier et al., fig. 9a (cup).
2007 Lonchocrinus sp.; Charbonnier et al., fig. 9b–c (second primibrachials).
Material. Pont des Étoiles: 2 cups. La Pouza: 8 cups, 4 s primibrachials. Chénier Ravine: 4 cups. La Clapouze: 68 cups, 9 s primibrachials.
Description. The cup has a cylindrical proximal and a widened upper part and it consists of radials with distinct interradial processes. The second primibrachials are axilllary, with a prominent spine-like process separating small facets for the secundibrachials (Fig. 12b). In the material from La Pouza are two axillaries with blunted tip curved inward (Fig. 12c).
Remarks. The species is most common at La Clapouze. It is easily distinguished from Eugeniacrinites cariophilites von Schlotheim by the shape of the second primibrachials and the different profile of the cup.
-
Family Phyllocrinidae Jaekel, 1907
-
Phyllocrinus d’Orbigny 1850
1871 Eugeniacrinus fenestratus Dumortier, p. 49, pl. 5, fig. 14–16 (cups).
1882 Phyllocrinus fenestratus (Dumortier); de Loriol, p. 106, pl. 17, figs. 3–9 (cups).
Material. The species has been found by the author only at La Clapouze where it is even more common than Lonchocrinus dumortieri. In fact, some rock samples are literally filled with the cups of this species. There are 96 isolated cups and several rock fragments containing approximately 35 cups. Dumortier (1871) also reported the species from La Pouza and Rians (Var), and de Loriol (1882) reported it from the area of Aix.
Description. The cups are circular in the lower part, increase in diameter towards the articular facets and are prolonged interradially into high processes of triangular section (see Hess & Messing 2011, fig. 89, f). The articular facets are deeply sunken between the processes. The facet to the column is small and deeply concave in most specimens. A specimen preserved on rock has exceptionally high processes and a rather large facet to the column (Fig. 12e). Variations include a rather broad cup with bulging radials and small facet to the column (Hess & Messing 2011, fig. 89, d–f), slender cups with small column facet, the most common form at La Clapouze; one of such specimens has a juvenile cup attached (Fig. 12d) and cups with processes curved more or less outwards (Dumortier 1871, pl. 5, fig. 14; de Loriol 1882, pl. 17, fig. 4).
Remarks. In the material from La Pouza sampled by A. S. Gale, no cup of this species has been found. This is rather surprising, given the report by Dumortier. Perhaps, Dumortier’s specimens came from parts of the outcrop rich in sponge remains, similar to the Clapouze site, whereas most of the present material is from the claystone beds.
-
Phyllocrinus colloti de Loriol 1882, Fig. 13a
Fig. 13 
Phyllocrinidae, Lower Bathonian, La Pouza (a–e) and Upper Bajocian-Lower Bathonian, Pont des Étoiles (g). a Phyllocrinus colloti de Loriol; cup with three fused columnals, a1 lateral, a2 lower (distal facet of third columnal), M10944. b–e Phyllocrinus voultensis n. sp.; b cup, holotype, M10854; b1 oblique lateral-distal view, b2 distal view, b3 proximal (aboral) view; c distal view of larger cup, M 10902; d oblique lateral view of cup with rather wide radials, paratype, M 10855; e lateral view of small cup with strongly bulging radials, M 10851. f Scutellacrinus tenuis n. g. n. sp.; cup, holotpe, M10839; f1 proximal (aboral) view, f2 oblique distal (upper) view, f3 lateral view. Scale bars 1 mm

1882 Phyllocrinus colloti de Loriol, p. 106, pl. 18, fig. 4–9.
2007 Phyllocrinus colloti de Loriol; Charbonnier et al., fig. 10b–c.
Material. Pont des Étoiles: 3 cups. La Pouza: 1 cup, Chénier Ravine: 4 cups (one with 9 and one with 10 columnals).
Description. This is the only cyrtocrinid from the area with at least one, but commonly several, columnals attached to the cup. The majority of specimens are from the Chénier Ravine locality, and it has not been found in the residues of the La Pouza clay beds. The cup is smooth and considerably higher than wide; it is circular in section and consists of commonly fused radials. The articular facets for the primibrachials are wide for a member of the genus, the interradial processes are strong at the base, moderately high and triangular in section. There is a gradual transition to the first columnal whose facet is rather wide, but is never exposed because of fusion. The cup in Fig. 13a is attached to three fused columnals. The column may have straight latera, but columnals may be slightly convex (Charbonnier et al. 2007, fig. 10b–c). Column facets have short marginal crenulae (Fig. 13a2).
Remarks. Charbonnier et al. (2007, fig. 10b–c) figured two specimens from Chénier, one with a short column and the other with a long one, similar to the material collected by the author. The specimens figured by de Loriol (de Loriol 1882, pl. 18, figs. 4–9) are from Rians (Var) and do not differ from those of the Ardèche area.
-
Phyllocrinus voultensis n. sp., Fig. 13b–e
Material. Pont des Étoiles: 3 cups, La Pouza: 14 cups.
Holotype. Cup, diameter and height 4 mm, M10854, Fig. 13b.
Etymology. After the type locality.
Type locality and horizon. Early Bathonian (Zigzag Zone), La Pouza, La Voulte-sur-Rhône (Ardèche, France).
Diagnosis. Cup small, pentalobate in section, with interradial furrows; facet to column sunken, with short marginal crenulae and wide axial canal; radial cavity narrow and deep; radials strongly bulging radially, with moderately high interradial projections, triangular in section; facets with small aboral ligament fossa and deep, circular muscle fossae.
Description. A number of small, low cups differ from other species of Phyllocrinus from the Ardèche. They have outward bulging radials, which are slightly keeled with deep interradial furrows in the smallest specimen (diameter 2 mm, height 1.6 mm; Fig. 13e), but are commonly rounded (Fig. 13b, d); the largest specimen is somewhat flattened (Fig. 13c). Sutures between the radials are distinct and sunken. The facets lie between triangular interradial projections. The facet to the unknown column is deeply sunken in the smallest specimen, larger and moderately sunken in the holotype, and nearly flush with the aboral side of the cup in the largest cup. The radial cavity is deep, extending to near the bottom.
Remarks. The present species is part of a series of small Phyllocrinus characterized by a nearly spherical cup with rather small interradial processes and bulging radials. The group includes P. gauthieri de Loriol (1882, pl. 17, fig. 2–3) from the Bathonian of Claps (Bouches-du-Rhône), P. alpinus (d’Orbigny) described by de Loriol (1882, p. 18, fig. 2) from the Oxfordian of Chaudon (Basses-Alpes) and P. gibbosus de Loriol (1882, pl. 18, fig. 3) from the Oxfordian of Crussol (Ardèche). Phyllocrinus voultensis n. sp. is closest to P. alpinus, known from a unique specimen with a diameter of 2.5 mm and a similar height (a height of 3.5 mm was given by de Loriol, but the figure shows a height similar to the diameter). The radials are keeled, and the cup is nearly pentagonal in outline. De Loriol’s specimen is also distinguished from P. voultensis n. sp. by a wider aboral ligament fossa.
-
Scutellacrinus n. g.
Diagnosis. Cup flat, shield or dish-like, composed of thin radials without bulges, radial cavity deep and wide, radial articular facets wide, separated by low, leaf-like interradial processes; facet to column small.
Etymology. After the shield- or dish-like cup.
Type species. Scutellacrinus tenuis n. sp.
Remarks. The cup combines the wide articular facets and interradial projections of a eugeniacrinitid with the generally slender build and small columnal facet of a phyllocrinid. The wide articular facets, not typical of phyllocrinids, obviously are a consequence of the dish-like geometry of the cup. The genus is tentatively assigned to the Phyllocrinidae.
-
Scutellacrinus tenuis n. sp., Fig. 13f
Material. Only the holotype is available.
Holotype. Cup, diameter 4.5 mm, height 2.7 mm, M10839
Etymology. After the thin radials.
Type locality and horizon. Late Bajocian/Early Bathonian (Parkinsoni/Zigzag Zones), Pont des Étoiles, La Voulte-sur-Rhône (Ardèche, France).
Diagnosis. See genus (monotypic).
Description. The cup is wide and low. The articular facets for the primibrachials occupy most of the width and extend into the cavity; the facets are almost entirely occupied by the large ellipitical muscle fossae, which are separated by a narrow notch. The aboral ligament is reduced to a small pit and interarticular ligament fossae appear to be absent. The interradial processes are leaf-like, with rounded tips; their sides are flanked by the muscle fossae. The wall of the radial cavity is evenly sloping and is nearly smooth (a larger crystal is seen in the cavity). The facet to the column is indistinct, but is small in any case.
Remarks. There is no cyrtocrinid which resembles this peculiar form.
-
Family Sclerocrinidae Jaekel, 1918
-
Cyrtocrinus Jaekel, 1891
-
Cyrtocrinus praenutans n. sp., Figs. 14, 15, 16d–e, 17–21, 24–25
Fig. 14 
Cyrtocrinus praenutans n. sp., Lower Bathonian, La Pouza. a Cup. holotype, M10845; a1 proximal, a2 lateral, a3 distal. b Lateral view of high cup with pit, M10828. c Oblique distal view of juvenile cup, M10941. d Proximal view of small cup, M 10942. e Distal view of juvenile cup, M 10904. f Lateral view of broken cup showing two radials with zygosynostosial facets, M 10901. g Oblique lateral view of isolated radial with pit on aboral side, M 19832. h Lateral view of cup attached to uppermost columnal with pits on radials and columnal, M 10899. i Broken cup and uppermost columnal with stunted columnal in aboral cavity, M 10835. j Distal view of tetramerous cup, M 10865. k Aboral view of radial with verrucose surface, M11000. Scale bars 1 mm
Fig. 15 
Cyrtocrinus praenutans n. sp., Lower Bathonian, La Pouza, first and second primibrachials. a Distal view of second primibrachial with one higher extension, M10896. b Second primibrachial, b1 proximal view, b2 distal view, M10858. c Proximal view of second primibrachial, M 10864. d Oblique proximal view of second primibrachial with pit, M 10905. e Distal view of compact second primibrachial, M10862. f Aboral view of fused first and second primibrachials, note shield-like upper part corresponding to second primibrachial and short lower part corresponding to first primibrachial, M 11032. g Proximal muscular facet of fused first and second primibrachials with verrucose surface, M 10908. h Distal view of second primibrachial with adorally united extensions, M10919. i Adoral view of first primibrachial with large articular facet and one side extended, M10894. j Adoral view of fused first and second primibrachials, this ossicle is much higher than Fig. 15f or g, M19893. k Adoral view of small first primibrachial, M10870. l First primibrachial, paratype, l1 adoral-proximal view, l2 aboral-distal view, M10861. Scale bars 1 mm
Fig. 16 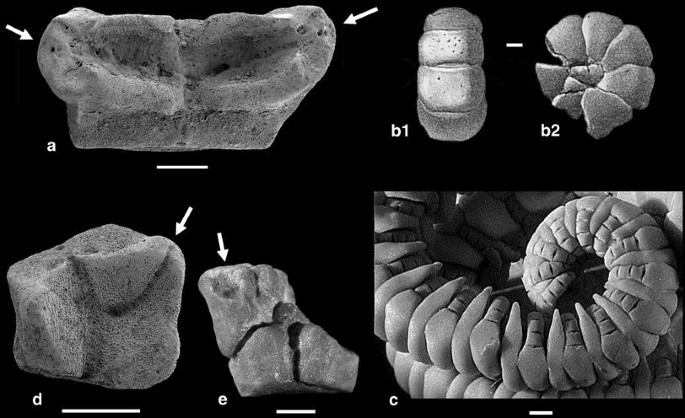
Pinnulation of cyrtocrinids. a Cyrtocrinus nutans (Goldfuss), Birmenstorf Member (Middle Oxfordian), Trimbach, Switzerland, M19836; adoral view to show the two pinnule sockets (bipinnulate condition). b Cyrtocrinus nutans (Goldfuss), Oxfordian, Streitberg, Bayerische Staatssammlung für Paläontologie und und historische Geologie München (Jaekel 1891); curled arm of rectangular brachials, b1 aboral view, b2 lateral view. c Living Neogymnocrinus richeri Bourseau et al., New Caledonia, M10980; specimen shows curled arm with pinnules on narrow, thickened part of brachials. d Cyrtocrinus praenutans n. sp., Lower Bathonian, La Pouza, paratype, M10848; adoral view with food grooves and pinnule socket (arrow), distal facet at upper side. e Cyrtocrinus praenutans n. sp., Lower Bathonian, La Pouza, M11033; adoral view of axillary second primibrachial with attached first secundibrachial. Arrows denote pinnule sockets. Scale bars 1 mm
Fig. 17 
Cyrtocrinus praenutans n. sp., Lower Bathonian, La Pouza, secundibrachials. a Secundibrachial, a1 adoral (pinnule socket to the right), a2 distal, M10846. b Distal facet of secundibrachial, paratype, pinnule socket is on extension at right, M10881. c Distal view of small asymmetric secundibrachial with verrucose surface, M10859. d Oblique lateral-distal view of block-like secundibrachial with pinnule socket on high side, M 10898. e Distal view of secundibrachial with high extension, pinnule socket is on low part at right, M10879. f Adoral view of secundibrachial with pit-like proximal muscle fossa facet at lower left, M10909. g Distal view of small secundibrachial with pronounced verrucose surface, M10834. h Adoral view of thin secundibrachial, M10869. i Synostosial facet of low weakly verrucose secundibrachial lacking pinnule socket, M10860. j Verrucose secundibrachial, j1 proximal synostosial facet, j2 distal muscular facet with pinnule socket, M10857. k Small, asymmetric thin secundibrachial, k1 adoral view, k2 upper facet, axial canal is below the food groove at right, M10995. Scale bars 1 mm, the short one only refers to Fig. 15e




1882 Eugeniacrinus nutans Goldfuss; de Loriol, p. 106, pl. 12, figs. 17–23 (cups, columnals).
1928 Eugeniacrinus voultensis Lissajous 1919, proposed by Sayn & Roman, p. 141, for large verrucose or granular cups with radials of equal size.
2007 Cyrtocrinus nutans (Goldfuss); Charbonnier et al., fig. 9d–h, (brachial, cups).
2007 Cyrtocrinus nutans var. voultensis; Charbonnier et al., fig. 9i (verrucose cup).
2007 Gammarocrinites compressus (Goldfuss); Charbonnier et al., fig. 9j (cup).
Material collected by the author from the four localities (not included in statistical analysis): 347 cups, 26 cups with pits, 28 cups with topmost columnal attached, 69 topmost columnals, 357 columnals, 13 columnals with pits, 37 attachment discs, 1 first primibrachial, 41 s primibrachials and 46 secundibrachials.
Material from La Pouza sampling (A. S. Gale, for details see Tables 2–4; Appendix 1, 2): 2,443 cups and radials (calculated as cups), 1,814 top columnals, 7,990 columnals, 1,069 attachment discs, 4,165 first primibrachials, 11,921 s (axillary) primibrachials and 58,744 secundibrachials.
Holotype. Cup, Fig. 14a, M10845.
Paratypes. Cup, Fig. 14b, M10828; secundibrachials Fig. 16d, M10990 and Fig. 17a, M10846.
Etymology. Precursor to nutans.
Type locality and horizon. Early Bathonian (Zigzag Zone), La Pouza near La Voulte-sur-Rhône (Ardèche, France).
Diagnosis. Cup low, smooth, rarely verrucose, circular and turban-like in most specimens, rarely somewhat higher and pentagonal with convex radials separated by furrows; cup nearly symmetric in most cases, facet to columnal not much displaced and angle between cup and column not more than approximately 20°. First and second primbrachials articulated by synostosis, exceptionally fused; first primibrachials may have lateral extension, second primibrachials not uncommonly with one or two adoral extensions, exceptionally united adorally. First pinnule on first secundibrachial. Secundibrachials never bipinnulate, the large majority with two muscular facets, rarely one facet synostosial and, exceptionally, two synostosial facets. Surface of brachials mostly smooth, but may be slightly to strongly verrucose; ornamentation increases from first to second primibrachials to secundibrachials, distal brachials more verrucose than proximal ones.
Description. Deepened analyses of some aspects are contained in the sections Ecological interactions of Cyrtocrinus praenutans n. sp., Reconstruction of Cyrtocrinus praenutans n. sp., and Classification of Cyrtocrinus praenutans n. sp. as influenced by ontogeny and variability.
Cups are low, circular to rounded pentagonal and rather symmetric in most specimens, with somewhat convex radials. Some large specimens are higher (Fig. 14b). Juvenile cups commonly have a pentagonal profile with notches between the radials (Fig. 14c–e). The surface is smooth and only exceptionally more or less verrucose or granular (Fig. 14k). The radials are joined by synostoses whose surfaces are ornamented by a pattern of dense crenulae (zygosynostosis) (Fig. 14f, g). The lower, proximal side of most cups is a wide and moderately deep bowl for reception of the topmost columnal. The distal, upper side has a mostly rather wide radial cavity sloping to the base of the cavity (Fig. 14f). The cavity may be sculptured into elevated interradial segments (Fig. 14a3, i, j), but in other specimens, such sculpturing is not developed (Fig. 14e, f). Radial articular facets are wide and extend to near the edge of the radials. The aboral ligament fossa is wide and shallow; the ligament pit oval and deep, and it may be small (Fig. 14a2) or wide (Fig. 14k). The transverse ridge is narrow, with a small axial canal (Fig. 14a2, k). The paired muscle fossae are circular and deep, with some structure and they are separated by a notch.
Topmost columnals vary greatly in shape, diameter and height (Fig. 19). Some specimens are preserved with a columnal still attached to the cup (Fig. 19k, m), and in such cases a small topmost columnal may be hidden in the aboral cavity of the cup. In a similar case, the topmost columnal is seen from the outside as a triangular, wedge-shaped element (Fig. 19l). Many specimens have topmost columnals contained in the cavity of the cup; they are low, nearly flat pyramids or cones seen from below (Figs. 18b, c, 19a, c1, n1) and they are articulated to the cup by irregular crenulae or ridges on the upper side (Fig. 19c2). However, the upper facet may also be flat (Fig. 19n2). A number of columnals is more or less oval; heights differ (Fig. 19e, g–i), and the decision as to which side is proximal must be based on the presence of irregular crenulae on the upper side and regular, stronger crenulae on the lower facet, articulated to the column. However, such decision is not possible with a columnal such as in Fig. 19i. Finally, a number of rather high, presumed top columnals have facets that are different at both ends and at an angle to each other (Fig. 19d, j). Again, the facet with regular, radiating crenulae at the cylindrical end is considered to be the lower.

Cyrtocrinus praenutans n. sp., Lower Bathonian, La Pouza. a–j, with pits of Oichnus paraboloides Bromley. a Lateral view of cup with pit surrounded by rim, M10830. b Proximal (lower) side of cup with stunted columnal, two deep pits are on the swollen radial at the right and shallow pits on the lower radial and between the radials at the left, M10829. c Proximal (lower) side of juvenile cup with stunted columnal and pit, M10829. d Aboral view of fused first and second primibrachials with pit extending over both ossicles, M 10897. e Aboral view of partly fused first and second primibrachials with large pit on second primibrachial, M 10880. f Proximal facet of second primibrachial with pit extending from the aboral side to the proximal facet, M10833. g Distal facet of secundibrachial with pit on the aboral side, pinnule socket at the upper right, M10907. h Aboral side of second primibrachial with pit, M 10831. i Proximal-aboral side of second primibrachial with three pits, M10884. j Columnal with large pit on latus, M10895. k Pit of unknown origin on latus of barrel-shaped columnal, flat pit with central canal occupies most of latus on one side (pit is to the right on Fig. 7k1), k1 upper facet, k2 latus with pit (hole in the upper part is due to weathering), M10993. Scale bars 1 mm

Topmost columnals of Cyrtocrinus praenutans n. sp., Lower Bathonian, La Pouza. a Proximal view of cup with low pentagonal columnal on thin plates in the aboral cavity of cup, two radials have pits, M10954. b Higher columnal, b1 upper facet with central depression, b2 lower facet with short strong crenulae, M10961. c Cone-shaped partly angular columnal, c1 distal facet, c2 proximal facet with irregular short crenulae, M10963. d High columnal with oval upper part, d1 proximal facet with wide depression, d2 lateral, d3 distal facet with irregular crenulae, M10965. e Aboral view of cup with bulging radials and oval columnal, M10956. f Upper facet of circular columnal with central depression surrounded by five sloping facets with additional depressions, lower facet with a few short crenulae, M 10847. g Oval columnal with pit at both narrow ends, M10910. h Low saddle-shaped oval columnal, h1 proximal facet with a few irregular crenulae, h2 lateral, h3 distal facet with long crenulae, M10966. i Saddle-shaped columnal, i1 proximal facet (distal facet is similar), i2 lateral, M10964. j Lateral view of high columnal with sloping proximal facet at an angle of approximately 45° to the cup, M 10967. k Aboral view of cup with columnal attached at an angle, lower facet of columnal with strong long crenulae, M10955. l Slightly asymmetric cup with triangular, wedge-shaped topmost columnal (arrow), M10991. m Nearly symmetric cup with attached high columnal (small topmost columnal may be hidden), M10992. n Small conical columnal, n1 lower facet, n2 upper facet with wide axial canal, M10999. Scale bars 1 mm
Attachment discs or holdfasts (Fig. 20). The majority of holdfasts are cone-shaped discs, with a concave to flat lower side. Some discs are root-like and probably anchored in soft sediment (Fig. 20g). Exceptionally, discs are attached to a foreign object such as a small oyster (Fig. 20e). Facets to the column vary in diameter and are deeply concave, with a strong rim that may show crenulae.

Attachment discs of Cyrtocrinus praenutans n. sp. with facet to columnal, Lower Bathonian, La Pouza. a Small disc, M10950. b Disc with wide facet to column, M10951. c Upper side of disc with three pits, M10996. d Flat disc with distinct crenulae, M10953. e Disc attached to small oyster, M10997. f Cone-shaped disc, M10958. g Large disc with root-like base, M10959. Scale bars 1 mm
Columnals (Fig. 21). Pluricolumnals are exceptional in the material and consist of only two ossicles (Fig. 21b). There is great variation in size, height, diameter, shape and articular facet. Columnals have smooth latera and are mostly more or less cylindrical, but may also be slightly convex or concave. In a given columnal, the width of lumen may be narrow at one end and wide at the other, with corresponding differences in crenulation (Fig. 21f). Given the commonly wide lumen of the attachment discs, it is assumed that a narrow lumen is on the proximal facet. Low, cylindrical or lens-like columnals seem to be proximal ones (Fig. 21e, l), and higher columnals may be distal. High, thin columnals (Fig. 21j, m) may be juvenile. Given all the shapes, it is surprising that the reconstructed animal should have only four columnals (see Table 4). Some small columnals are branched (see Appendix 2). They either branch symmetrically, V-like (Fig. 23b) or a branch originates from the side (Fig. 23c). The facets of these columnals do not differ from facets of other small columnals. The riddle of axillary columns in Cyclocrinus, millericrinids and cyrtocrinids has been discussed by Hess (2008, p. 475) and the present material adds still another example.

Columnals of Cyrtocrinus praenutans n. sp., Lower Bathonian, La Pouza. a High spindle-shaped columnal, a1 upper facet, a2 lateral, a3 lower facet, M 10968. b Pluricolumnal of two convex ossicles, b1 upper facet, b2 lateral, M10969. c Concave columnal, c1 upper facet (lower facet is similar), c2 lateral, M10970. d Cylindrical columnal, d1 upper facet with strong irregular crenulae (lower facet is similar), d2 lateral, M10971. e Convex columnal, e1 presumed proximal concave facet, e2 lateral, e3 presumed distal facet, M10972. f High columnal, f1 presumed proximal facet, f2 lateral, f3 presumed distal facet with wide lumen and strong crenulae, M10973. g High concave columnal, g1 presumed proximal facet, g2 lateral, g3 presumed distal facet with wide lumen and weak crenulae, M10974. h Columnal with irregular crenulae and narrow lumen, h1 upper facet, h2 lateral, h3 lower facet, M10975. i Slightly conical columnal with similar facets, i1 upper facet, i2 lateral, i3 lower facet, M10976. j High thin columnal, j1 upper facet, j2 lateral, M10977. k Convex columnal, k1 upper facet (lower is similar), k2 lateral, M10978. l Lens-like columnal, l1 upper facet, l2 lower facet, M10960. m Small thin columnal, M10998. Scale bars 1 mm
First primibrachials (Figs. 15i–l, 24i–o). These ossicles articulate to the cup, their proximal facet matching the facet of the cup. Typically, first primibrachials have proximal facets with pronounced muscle fossae (Fig. 15k, l1) and are relatively thin, with a pronounced adoral furrow. Their outline is mostly trapezoidal, but one side may be extended (Fig. 15i). The distal facet is synostosial (Figs. 15l2, 24i–k, m–o), rarely cryptosyzygial (Fig. 24l) and crescent shaped. In some cases, first primibrachials are fused with second primibrachials. Such a fused pair may vary from high (Fig. 15j) to lower (Fig. 25g) to low (Fig. 18d–e); one fused pair has a low proximal (=first primibrachial) part (Fig. 15f).
Second primibrachials (Figs. 15a–h, 24a–h, 25). These distinctive ossicles with a proximal synostosial facet and two distal muscular facets occur in a variety of shapes. The majority are low to high trapezoidal in outline (Figs. 15g, 24a–h, 25e–f). More remarkable are sturdy ossicles with unilateral or paired adoral extensions of variable shape and thickness, and adoral furrows are mostly deep (Figs. 15a–e, 18f). Exceptionally, the processes are completely fused, leaving only a narrow passage for the food groove (Fig. 15h). The figured ossicle resembles second primibrachials of the Oxfordian Pilocrinus moussoni (Desor) (see Hess & Messing 2011, fig. 92g–h).
Secundibrachials (Figs. 16d–e, 17, 18g). Secundibrachials are by far the most common fossils in the La Pouza samples, and they occur in a large variety of sizes and shapes. Because no axillary secundibrachial with a proximal muscular facet has been found, the arms were unbranched after the axillary second primibrachial, and the animal had only ten arms. Proximal brachials are more or less rectangular and massive (Fig. 17a, b, d). They bear only one pinnule socket, situated on the thickened part. This socket is supplied with a branch of the axial food groove (Figs. 16d, 17a, f, h). A first secundibrachial still attached to the second primibrachial has a deep pinnule socket near the distal margin (Fig. 16e). The following pinnule sockets are on the lateral margin, as in the extant Neogymnocrinus richeri (Fig. 16c). No pinnulars were isolated from the washings, but it may be assumed that they were similar to the extant example. A number of secundibrachials have one side extended club-like (Fig. 17e, pinnule socket is on low part at right). The interbrachial facets have small, distinct muscle fossae below a notch for the food groove; the muscle fossae are separated by the axial canal and the transverse ridge from the narrow aboral ligament fossa with its pit. Most of the proximal secundibrachials are smooth, but distal ossicles are increasingly verrucose (Fig. 17c, g). Among the material is a certain percentage of small, thin brachials with the food groove displaced to one side (Fig. 17k). Muscle fossae are small and rather indistinct. It is assumed that such ossicles are from distal parts of the arm. An even smaller percentage of secundibrachials have one or both facets synostosial (Fig. 17i, j) (see Appendix 1). Brachials with synostoses on both facets are thin and lack a pinnule socket. Their position in the arm is unclear. Secundibrachials with synostosial facets do not normally occur in cyrtocrinids where facets are muscular, although synostosial facets of a cyrtocrinid species from the lower Jurassic have been reported by Hess (2006, pl. 6, fig. 16–17).
Remarks. This species dominates the La Pouza fauna, but also occurs in the Bajocian/Bathonian of Pont des Étoiles, the Bathonian of La Clapouze and the Lower Callovien of the Chénier Ravine. It was previously assigned to the Middle Oxfordian C. nutans. In fact, some cups closely resemble those from the Oxfordian. However, the large amount of material now available from La Pouza demonstrates that there are significant differences between the two species separated by a wide stratigraphical gap. Cyrtocrinus nutans was extensively described by Quenstedt (1876) who figured a large number of elements; unfortunately, his figures are so small that details cannot be recognized in many cases. Quenstedt figured attachment discs, each with wide lumen (1876, pl. 105, figs. 144–155), a feature comparable to the discs of C. praenutans that are, however, mostly cone- or dish-like and only rarely root-like. Branched columnals are also recognized in both species, as are columnals of variable shape and length. The cups of C. nutans are commonly more oblique and the radial articular facets may be at a right angle to the column axis. In such cases, the cup may be fused with the top columnal. Such extreme features are unknown from C. praenutans where cup variability is much less. One might say that C. praenutans has a lower degree of crookedness. Of special relevance to the establishment of a new species for the Ardèche material are descriptions of bipinnulate secundibrachials in C. nutans (Quenstedt 1876, pl. 106, figs. 62, 63; p. 434–435). The rectangular aboral profile of the brachials on a curled arm (Fig. 16b) also supports the bipinnulate condition of the Oxfordian species. Well-preserved material from the Middle Oxfordian Birmenstorf Member of northern Switzerland shows this in the necessary detail (Fig. 16a). Incidentally, de Loriol (1889, p. 560; pl. 229, figs. 10–11) described bipinnulate secundibrachials from the Oxfordian of Le Pontet near Saint-Claude (Jura), and thought that they belonged to his Gymnocrinus moeschi (=adorally enclosed second primibrachials of Pilocrinus moussoni [Desor]) (see Hess & Messing 2011, p. 189). Cyrtocrinus nutans also occurs at Saint-Claude. The arms of C. praenutans n. sp. may have resembled those of the living Neogymnocrinus richeri (Bourseau, Améziane-Cominardi, Avocat & Roux 1991) (Fig. 16c), more or less rectangular brachials proximally, and cuneate brachials distally. Cups of the present species from the Lower Callovian of the Chénier Ravine have been figured by Charbonnier et al. (2007), in part under different names. This is discussed in a following section.
-
Superfamily Plicatocrinoidea Zittel, 1879
-
Family Tetracrinidae Nicosia, 1991
-
Praetetracrinus Jäger, 1995
-
Praetetracrinus bathonicus n. sp., Fig. 22
Fig. 22 
Praetetracrinus bathonicus n. sp., Lower Bathonian, La Pouza. a Radial, holotype; a1 aboral, a2 adoral, M10875. b Adoral view of proximal part of radial with two pairs of muscle fossae, M10866. c Radial articular facet, M10871. d Aboral view of small verrucose radial, M10892. e Adoral view of small low radial, M10868. f Proximal (lower) facet of pentamerous basal circlet, M10874. g Proximal (lower) facet of tetramerous basal circlet, M10873. h Tetramerous basal circlet, M10877, paratype; h1 upper (distal) view with facets to radials, h2 oblique proximal-lateral view. i Facet of columnal, M10867. Scale bar 1 mm

Material. Basal elements: 24 (one fived sided, 23 four sided); 56 radials (37 intact, 18 partly broken), 3 columnals.
Holotype. Radial, largest width 3 mm, height 2.5 mm, M19875, Fig. 22a.
Paratype. Tetramerous basal element, diameter 2.5 mm, M10877, Fig. 22h.
Etymology. After the stratigraphic occurrence.
Type locality and horizon. Early Bathonian (Zigzag Zone), La Pouza, La Voulte-sur-Rhône (Ardèche, France).
Diagnosis. Basal element tetramerous, exceptionally pentamerous, lower side slightly concave with radial crenulae mostly in groups, upper side with moderately deep cavity and small facets to radials at the angles with paired axial canals. Radials thin walled, commonly high, with smooth, slightly keeled aboral side, rarely lower and granular aborally; base narrow, facet for primibrachial wide, leading to conical shape of cup with wide radial cavity; radial articular facet crescent shaped, raised into short process at each end, aboral ligament fossa with oval pit, transverse ridge distinct, muscle fossae wide and extending onto the adoral side with lower rim. Columnals circular and facets with four crenulae (corresponding to tetramerous symmetry).
Description. The basal elements are mostly tetramerous, with only one pentamerous specimen. Height is roughly equal to diameter. The facet to the column is circular and concave, with mostly four groups of two to three distinct crenulae. The upper facet is quadratic or pentagonal in outline (depending on the symmetry) and the rim is widened at the corners where paired nerve canals are visible. Radials vary in shape and height. Most are high, thin and aborally smooth, but some are nearly triangular with granular aboral surface (Fig. 22d, e). Most radials are slightly keeled aborally. The radial articular facets have wide, oval muscle fossae with adoral rim and separated by rimmed notch on the inner, adoral side. The muscle fossae are separated by a distinct transverse ridge from the small aboral part with an oval ligament fossa. A radial with apparently two pairs of muscle fossae is shown in Fig. 22b.
Remarks. The elements in question belong to the Plicatocrinoidea Zittel. This superfamily includes two families with a distinct element of fused basals and a variable number of commonly high radials, Plicatocrinidae Zittel and Tetracrinidae Nicosia. The present elements are assigned to the genus Praetetracrinus based on the tetramerous symmetry of the stout basal element. Praetetracrinus bathonicus n. sp. resembles the Early Jurassic P. inornatus (Simms 1989) but has thinner and higher radials; in addition, the basal element is higher and angular. From the Toarcian/Aalenian of Germany, Jäger (1995, pl. 6, figs. 9–12) figured several elements with characters closely resembling those of P. bathonicus: a slender, high radial (loc. cit., fig. 12), a columnal (loc. cit. fig. 9), and two basal elements (loc. cit. figs. 10–11). Jäger classified the ossicles as either P. kutscheri Jäger or P. inornatus (Simms). The radials of the Toarcian/Aalenian P. doreckae Jäger (1995) have straight to sligthly concave upper edges and are not keeled aborally. The basal circlet of the Pliensbachian Plicatocrinus sulzkirchenensis Jäger (1993) is penta- to heptamerous with high and narrow radials. In comparison with the Pliensbachian-Toarcian Quenstedticrinus deslongchampsi (de Loriol) (de Loriol 1882, pl. 10; Hess 2006, pl. 6, fig. 1), P. bathonicus n. sp. has thinner and adorally unsculptured radials. Quenstedticrinus Klikushin belongs to a separate family, Quenstedticrinidae Klikushin, 1987, characterized by a cryptic basal circlet within the radial circlet. Plicatocrinus hexagonus Münster from the Middle Oxfordian sponge facies of Germany and Switzerland is mostly hexamerous with a thin, shallow basal element; but the similarly keeled and thin radials resemble those of the present species. Unfortunately, no brachials or pinnulars can be assigned to P. bathonicus n. sp. with certainty, preventing further elaboration of relationships. In contrast to Tetracrinus, the radials of Praetetracrinus and Sacariacrinus are high, resembling those of Plicatocrinus. Jäger (1995), not considering Nicosia’s (1991) establishment of the family Tetracrinidae, placed both Tetracrinus and Praetetracrinus in the family Plicatocrinidae. Thus, Jurassic Plicatocrinoidea appear to constitute a heterogeneous, presumably paraphyletic group, with P. bathonicus n. sp. more closely related to Early Jurassic than to Late Jurassic forms.
-
Suborder Holopodina Arendt, 1974
-
Family Eudesicrinidae Bather, 1899
-
Dolichocrinus de Loriol, 1891
-
Dolichocrinus aberrans (de Loriol 1882), Fig. 23a
Fig. 23 
Various specimens from the Lower Bathonian of La Pouza. a Broken cup of Dolichocrinus aberrans (de Loriol), M10918. b, c Branched columnals of Cyrtocrinus praenutans n. sp.; b columnal with symmetric branching, M10903; c columnal with lateral branch, M10979, c1 facets, c2 lateral view.d Columnal of Cyclocrinus rugosus d’Orbigny, M10926, d1 facet, d2 lateral, M10926. e Columnal of Apiocrinites sp., e1 facet, e2 lateral, M10925. Scale bars 1 mm

1882 Eugeniacrinus aberrans de Loriol, p. 148, pl. 15, figs. 4–5
1891 Dolichocrinus aberrans (de Loriol); de Loriol, p. 131; pl. 24, figs. 3–4
1891 Tetanocrinus aberrans (de Loriol); Jaekel, 1891, p. 628, fig. 15
2007 Dolichocrinus aberrans (de Loriol); Charbonnier et al., fig. 10a
2011 Dolichocrinus aberrans (de Loriol); Hess & Messing, 2011, fig. 101, 3
Material. The author collected eight cups from La Pouza and one cup from Chénier Ravine. The La Pouza sampling (A. S. Gale) furnished 22 cups, all of them more or less broken. Seventeen remains have radial facets preserved, 4 are only parts of the columnal element, and 1 shows facets to basals (see Hess & Messing 2011, fig. 101, 3b). The best preserved cup is 10 mm high, with a radial circlet diameter of 4 mm at the top and a diameter of the column part of 2.3 mm. The largest cup with only a short preserved column part has a diameter of 4.3 mm at the top; the smallest cup has a diameter of 2 mm (column part diameter 1 mm).
Description. The disarticulated or partly broken specimens reveal a wide axial canal between the prolonged, column-like radials (Fig. 23a). Except for size, variability is low between the specimens; radial articular facets are perpendicular to the axis or at only low angle to the column-like part. In the material from the La Pouza claystone beds, no specimen with an occluded radial has been found (Hess & Messing 2011, fig. 101, 3c; collected at La Pouza) and such morphology seems exceptional.
Remarks. The material isolated from the La Pouza claystone beds includes two single radials, nine radial pairs and three radial triplets (see also de Loriol 1882, pl. 15, fig. 6), as opposed to nine articulated specimens (8 with radial articular facets, 1 column part with facets to 5 basals). Such preservation is similar to that of Cyrtocrinus praenutans n. sp. where isolated radials or disarticulated parts of the cup are also common.
-
Incerti ordinis
-
Family Cyclocrinidae Sieverts-Doreck, 1953
-
Cyclocrinus d’Orbigny 1850
1871 Millericrinus; Dumortier, p. 46, pl. 5, figs. 7–11.
1886 Cyclocrinus macrocephalus (Quenstedt); de Loriol, p. 18, pl. 126, figs. 4–9.
2008 Cyclocrinus rugosus d’Orbigny; Hess, p. 465, text-figs. 1, 2; pl. 1, 2.
Material. Pont des Étoiles: 4 columnals; La Pouza; 34 columnals and 1 pluricolumnal of 2 columnals; Chénier Ravine: 4 columnals; La Clapouze: 3 columnals.
Description. The columnals are cylindrical, with smooth latera and are relatively high; the largest columnal from La Pouza has a diameter of 11 mm and a height of 6 mm; the smallest from this locality has a diameter of 2 mm with similar height. The facets are characteristic of the species. The facet in Fig. 23d1 is representative of the Ardèche material. It has rows of small tubercles radiating from the lumen; in between are larger tubercles and short crenulae or irregular tubercles along the margin.
Remarks. Dumortier (1871, pl. 5, fig. 7–11) figured a small columnal from La Pouza, a larger pluricolumnal of three and a large single columnal with a diameter of 18 mm. The facet of the small columnal (Dumortier 1871, fig. 8) has radial crenulae and tubercles; the facet of the large columnal (Fig. 11) has more or less radial and irregular, vermicose crenulae that extend from the lumen to near the margin. Such differences in ornamentation are common in this species, even in columnals from the same locality (Hess 2008).
Ecological interactions of Cyrtocrinus praenutans n. sp.
The material includes quite a number of ossicles with distinctive pits (Table 3). These are restricted to ossicles of Cyrtocrinus praenutans n. sp. and have been noted before (Ausich & Simms 1999, fig. 70, unnamed; Charbonnier et al. 2007, fig. 9h, as Tremichnus Brett on p. 228). Oval to circular non-penetrative pits have been named Oichnus paraboloides Bromley 1981 (p. 195), and the animal responsible is unknown. Such traces also occur in Palaeozoic crinoids (see, e.g. Donovan et al. 2010, pl. 31, fig. 1, on a crown of Sagenocrinites expansus [Phillips]). The highest percentage of pits in the Ardèche material is on cups, followed by attachment discs and elements close to the cup (topmost columnals and primibrachials). Attachment discs have more pits than columnals, perhaps because of their broader surface for epizoan attachment. Secundibrachials seem to be a less favourable substrate for the pit-forming epizoan. With possibly one or two exceptions, the pits are restricted to the aboral (outer) surface of the ossicles and do not reach the food grooves. A number of ossicles have multiple pits (Figs. 18b, d, i, 19a, 20c). Some fused first and second primibrachials carry pits that extend over both parts (Figs. 18d, 25g). Pits on cups are commonly accompanied by increased skeletal growth leading to swelling (Figs. 18a–b, 19a); such swelling is less common on brachials (Fig. 18f, i). On most brachials, columnals and attachment discs, and also on some cups or radials, pits are plain (Figs. 14b, g, 18a–j, 20c). A columnal has a large shallow depression of unknown origin on the latus; the depression has a convex surface with two small pits (Fig. 18k).
Reconstruction of Cyrtocrinus praenutans n. sp. (Table 4)
The number of top columnals, but especially of second primibrachials, corresponds rather well to the number of cups. The hypothetical animal would on average have only four columnals (including a mostly distinct topmost columnal). In view of the large variability of column shapes, this number seems to be rather low. It is true that the living Neogymnocrinus richeri Bourseau et al. (see Bourseau et al. 1991; Hess & Messing 2011) has only two columnals, but these are similar in shape, although not in length. First primibrachials number only approximately a third of second primibrachials. This is difficult to explain because the ossicles are easily recognized. First primibrachials are lower and blade-like in comparison with the mostly sturdy and angular second primibrachials. Thus, they may have been more susceptible to winnowing. However, such a sorting process would also have included the ossicles from the fine fraction (see Appendix 2), where the first and second primibrachials are equal in number. The number of secundibrachials seems too small for a cyrtocrinid of this type, with brachial morphologies differing along the arms.
Classification of Cyrtocrinus praenutans n. sp. as influenced by ontogeny and variability
The rich material of Cyrtocrinus praenutans n. sp., the only species of the genus found at La Pouza, invites a detailed analysis of variabiliy and ontogenetic changes and their influence on classification. All ossicle types are in principle available, that is attachment discs, columnals, topmost columnals, cups, first and second primibrachials, and secundibrachials. Apart from cups, ossicles most useful for an analysis are first, but especially second, primibrachials for the following reasons: (1) have a constant number of 5 in relation to the cup, (2) are easily recognized and sturdy, and (3) have a position at the base of the arms. First primibrachials, although directly attached to the cup, are impoverished in relation to cups (Table 4). Secundibrachials are considered less suitable because of a higher degree of variability in shape and ornamentation and the unknown arm length. In addition, the decision of whether a small brachial belonged to a juvenile animal or was on the distal part of the arm is not always straightforward.
A number of representative cups and primibrachials with variable shapes have been assembled in Fig. 24, reproduced at the same magnification. As discussed earlier, larger cups, presumably from adult individuals, are low, circular and turban-like, with convex radials separated by furrows (Figs. 14a, 18a, b, 19a, e, k, m). One modification is a higher cup (Fig. 14b). A similar cup is preserved in the Gevrey collection of the Université Joseph Fournier of Grenoble (UJF-ID.12088, Bathonian, Rompon) and is labelled Phyllocrinus gibbosus. However, P. gibbosus de Loriol (1882, p. 173, pl.18, fig. 3) from the Oxfordian of Crussol is a true Phyllocrinus species, with a narrow facet to the column and radial articular facets sunken between prominent interradial processes. In contrast, the Grenoble specimen has a wide, somewhat asymmetrical and deeply sunken facet to the column, and interradial processes are absent. Small cups commonly have more straight-sided aboral facets with deeper sutures, giving them a notched pentagonal profile as illustrated in Figs. 14c–e, 18c. Articular facets are also more prominent and relatively larger in relation to cup size (Figs. 14c, 24q. Charbonnier et al. (2007, fig. 9j) designated a smooth cup with rather narrow radial cavity from the Lower Callovian of the Chénier Ravine as Gammarocrinites compressus. However, the width of the radial cavity is not a distinctive character and larger cups from La Pouza commonly have narrower cavities (Figs. 14a3; 24p) than smaller ones (Figs. 14e, 24q).

Cyrtocrinus praenutans n. sp., Lower Bathonian, Bed 1, La Pouza. Series of cups (p, q, r), first primibrachials (i–o) and second primibrachials (a–h) to illustrate differences in size, shape, and ornamentation. Cups are shown in side view; in contrast to Fig. 15, primibrachials are shown in aboral view with synostosial facets exposed; first primibrachials are oriented with distal side upward mounted on muscular facets, second primibrachials show adoral facet facing downward. Asterisks indicate position of hidden proximal muscular facet articulating to radial. a M11022. b M11009. c 11020. d M11013, note narrow proximal synostosial facet. e M11007. f M11021. g M11018. h M11008. i M11030. j M11029. k M11025. l M11028, note cryptosyzygial distal facet. m M11031. n M11026. o M11027, note concave central part of facet. p M11002. q M11006. r M11001. Scale bar 1 mm
First primibrachials vary considerably in height and thickness (Fig. 24j–o). Distal articular facets to the second primibrachials are commonly crescent shaped and may vary in size; a range of sizes is illustrated in Figs. 24j, k, i, and n; occasionally the synostosial facet has fine marginal crenulae and resembles a cryptosyzygy (Fig. 24l). The facets cover the entire surface, but are rarely restricted to the middle part (Fig. 24o).
Second primibrachials mirror the first primibrachials in their variability (Fig. 24a–h). They range from thin and high (Figs. 24a–b, 25a–c, f), nearly trapezoidal in outline, to low triangular (Fig. 24d). Ossicles and, thus, their facets may be wide (Fig. 24f–h) or narrow (Fig. 24a–c). The aboral side may be straight (Fig. 24a–c), slightly convex (Fig. 24g) or slightly concave (Fig. 24e, h). Massive second primibrachials also vary in width, with corresponding elliptical or crescent-shaped proximal facets (Fig. 25d, e). Further examples are illustrated in Fig. 15 where primibrachials are figured in different positions.

Cyrtocrinus praenutans n. sp., Lower Bathonian, Bed 1, La Pouza. Series of second primibrachials (a–e) and fused first and second primibrachials f–g) to illustrate differences in size, shape, and ornamentation. Ossicles are oriented with distal side upward mounted on adoral side (a–c, e–g) or muscular facets (d). a M11023. b M11015, thin ossicle. c M11016, proximal facet narrow. d M11019, thick adoral extensions. e M11017, narrower, blunted oral extensions. f M11003, moderately high pair of fused first and second primibrachials, muscular facet to radial narrow. g M11004, rather thin pair of fused first and second primibrachials with suture still visible, note pit extending on both aboral facets. Scale bars 1 mm
Variability in ornamentation of the exterior (aboral) side of primibrachials is illustrated in Fig. 25, which also includes an ornamented fused first and second primibrachial with a pit extending on both ossicle parts (Fig. 25g). Granulation varies from a few scattered granules (Fig. 25g) to a more or less dense cover (Figs. 17g, 24c, 25f), making this character unsuitable for specific distinction. As listed in Appendix 1, attachment discs and columnals are never verrucose and cups are only rare (Fig. 14k). At La Pouza Bed 1, approximately 4 % of first primibrachials, 13 % of second primibrachials and 48 % of secundibrachials are verrucose, in particular the more distal ones (Fig. 17c, g). Thus, granulation increases from the cup to the more distal secundibrachials.
From the Chénier Ravine, Charbonnier et al. (2007, fig. 9d, i) figured a verrucose brachial as Cyrtocrinus nutans and a verrucose cup as C. nutans var. voultensis (Sayn & Roman 1928). While granulation may be more prominent in specimens from the Lower Callovian, a specific distinction based on this character is not feasible. Variability in cups of Cyrtocrinus nutans from the Middle Oxfordian of the Chalch Quarry (Holderbank) was discussed by Hess & Spichiger (2001, p. 490). They noted great variation in height, ornamentation and in the ratio of radial cavity diameter to cup diameter; the variation between 80 and 20 % corresponds to Quenstedt’s (1876) “subspecies” nutans apertus and nutans opertus. Previous assignment of ossicles with verrucose surface to Sclerocrinus (=Gammarocrinites) compressus was rejected by Hess & Spichiger, and brachials and cups ornamented by granules were considered part of intraspecific variation (see also Hess 2004 and Hess & Messing 2011, p. 188). Such treatment is supported by the present material. Verrucose specimens, designated by Goldfuss (1826–1833) as Eugeniacrinites compressus, were assigned by Jaekel (1891, p. 626) to his genus Sclerocrinus and by Rasmussen (Rasmussen 1978, p. T831) to Gammarocrinites Quenstedt. However, these may be merely variants or ecophenotypes of one species. The material from the Birmenstorf Member of Switzerland includes specimens with radial articular facet at a right angle to the column axis caused by either a triangular or a bent top columnal; the radials and the cup also exhibit a tendency to fuse with the top columnal. Such extremes are not seen in the Ardèche material.
Palaeoecology of the La Pouza site
Charbonnier et al. (2007, p. 231) postulated that the crinoids from the Bathonian exposure of La Pouza lived on a steep slope at a depth of about 150 m and it may be assumed that this is also true of the limited La Clapouze locality. According to these authors, the Lower Callovian crinoids and sponges of the Chénier Ravine lived on a carbonate platform and the adjoining slope in the upper bathyal zone at a water depth exceeding 200 m. Here, the submarine topography, with steep blocks and a fault-controlled escarpment, facilitated local sedimentary slides leading to autochthonous or parautochthonous burial of sponges and crinoids. These Middle Jurassic faunas are remarkable for the presence of a variety of phyllocrinids, and the first species of Eugeniacrinitidae (Lonchocrinus) and Sclerocrinidae (Cyrtocrinus); in contrast, stalkless members of Holopodina, so prominent in the Lower Jurassic, are entirely absent. This may have to do with the abundance of sponges that prevented colonization by crinoids occupying the lowest tier. As discussed above, living conditions of the Bathonian La Pouza fauna seem to have been similar to those of the Middle Oxfordian Birmenstorf Member. Both are diverse, essentially hardground faunas, living in moderately deep water.
Status of the Ardèche crinoid fauna
The Middle Jurassic Ardèche fauna bridges the gap between the rich Early Jurassic faunas from Arzo and Turkey (Hess 2006, table 1), England and Germany (Simms 1989; Jäger 1993, 1995), and the Late Jurassic (Middle Oxfordian) faunas from Germany and Switzerland (Quenstedt 1876; de Loriol 1877–79; Hess 1975; Hess & Spichiger 2001). In the Early Jurassic of Arzo, a surprising morphological variety of cyrtocrinids occurs. Members of the family Phyllocrinidae still possessed basals (Nerocrinus Manni & Nicosia, Ticinocrinus Hess); Eudesicrinidae include Eudesicrinus de Loriol and Dinardocrinus Manni & Nicosia with thick-walled cups, prolonged and column-like in the latter genus; Cotyledermatidae are represented by a species of Cotylederma Quenstedt with wide, thin cup attached to the substrate by a basal element; Plicatocrinidae are present with Sacariacrinus Nicosia, Quenstedticrinidae with two species of Quenstedticrinus Klikushin, and Tetracrinidae with Tetracrinus Münster, Bilecicrinus Manni & Nicosia and Arzocrinus Hess. Sclerocrinidae are documented by secundibrachials of “Gymnocrinus” sp.; Fusicrinus Hess and Castaneacrinus Hess are of uncertain familial assignment.
From the plethora of Early Jurassic forms, cyrtocrinids evolved in the Middle Jurassic into species more closely resembling those from the Late Jurassic. Phyllocrinidae now include only forms that lack basals and have columns with reduced diameter (Phyllocrinus fenestratus, P.colloti, P. voultensis n. sp.). The peculiar Scutellacrinus tenuis n. g. n., sp. may also belong in this group. However, Plicatocrinoida are represented by Praetetracrinus bathonicus n. sp., a form with characters reminiscent of Toarcian/Aalenian species of the genus. The eudesicrinid Dolichocrinus aberrans with its elongate cup has replaced Dinardocrinus tiburtinus Manni & Nicosia. Dinardocrinus has radials of extremely different length, and a remnant of this situation is seen in a cup of Dolichocrinus with occluded radial (Hess & Messing 2011, fig. 101, 3c). Cotyledermatids with wide cup attached via basal element to the substrate are absent and this group is restricted to the Early Jurassic. The Ardèche faunas are the first to include representatives of the Eugeniacrinitidae (Lonchocrinus) and Sclerocrinidae (Cyrtocrinus preanutans n. sp.). Species of these families are among the dominant cyrtocrinids in the Late Jurassic and Early Cretaceous, and this successful group has survived in the living sclerocrinid Neogymnocrinus Hess (see Hess & Messing 2011). Cyrtocrinus praenutans n. sp shares a number of characters with the Late Jurassic C. nutans, making it a perfect precursor to the latter.
A Middle Jurassic crinoid radiation is also seen in the comatulids, represented by five species, three of them new. Together with the Middle Jurassic Antedon (=Palaeocomaster) schlumbergeri and Antedon (=Andymetra) ladoixensis described by de Loriol (1889), there are now seven species in four genera. To these should be added three species of Semiometra, one from the Middle Jurassic of England, one from the Oxfordian of France and Poland, and one from the Oxfordian of Germany, see Radwanska (2007), Helm et al. (2003). From the Early Jurassic only three species have been described, belonging in two genera: Paracomatula liasica Hess, Palaeocomaster morierei (de Loriol) [possibly including P. caraboeufi (de Loriol, 1888)] from the Pliensbachian, and Palaeocomaster styriacus Kristan-Tollmann (1988) from the Hettangian (Hess 2006, Hess & Messing 2011). Palaeocomaster is the first comatulid proper with a centrodorsal consisting of a single piece that displays tube-like cirrus sockets. The peculiar Singillatimetra with only a single centrodorsal disc, which resembles the multiple discs of true Paracomatula, may be a remnant of Paracomatula that reached its peak in the splendid Bajocian P. helvetica Hess (Hess & Messing 2011).
Stalked isocrinids are represented in the Ardèche by several forms. The rare Pentacrinites ausichi n. sp. appears to be alien to the fauna because species of this genus mostly form beds or lenses of intact specimens (Hess 1999). More common are Balanocrinus with three species and Isocrinus with two species. The presence, at Pont des Étoiles, of three distinct species of Balanocrinus in the presence of the dominating isocrinid Isocrinus nicoleti seems unusual. I. nicoleti is considerably larger than the Balanocrinus species and, thus, would have occupied the highest tier of the crinoid fauna. However, the remains may not have come from a defined location because of its parautochthonous deposition. All three genera of Isocrinida are known from the Lower Jurassic and two persist into the Lower Cretaceous (Hess & Gale 2010). The uncommon Cyclocrinus rugosus and Apiocrinites sp. have a similar distribution.
Conclusions
The results demonstrate that the Middle Jurassic crinoids from the Ardèche constitute one of the important and diverse Mesozoic crinoid faunas. Some forms bridge the gap between the Early Jurassic and the Late Jurassic hardground faunas rich in cyrtocrinids. Cyrtocrinus praenutans n. sp., a form similar to Cyrtocrinus nutans from the Oxfordian, is described as a separate species despite overlapping phenotypic variability of cups and columnals. Pathological deformations on all types of ossicles of Cyrtocrinus praenutans n. sp. are ascribed to the epizoan commensal that created Oichnus paraboloides Bromley. Different species are dominant at the different Bathonian localities, Cyrtocrinus praenutans n. sp. at La Pouza, and Phyllocrinus fenestratus (Dumortier) and Lonchocrinus dumortieri (de Loriol) at La Clapouze. Preservation and rock formation of the Upper Bajocian–Lower Bathonian Isocrinus nicoleti (Thurmann) at Pont des Étoiles suggests that this form lived in rather shallow and turbulent water. The dominance of cyrtocrinids and the presence of all growth stages and parts of the skeleton at La Pouza and La Clapouze indicate a moderately deep palaeoenvironment and limited transport of the ossicles. The diversity of these Middle Jurassic faunas is not reached until the Middle Oxfordian sponge reef facies of southern Germany and Switzerland.
References
Alméras, Y., & Elmi, S. (1996). Le genre Cymathorhynchia Buckman (Brachiopoda, Rhynchonellacea) dans le Bajocien-Bathonian de la bordure vivaro-cévénole (Bassin du Sud-Est). Cadre stratigraphique et paléogéographique. Beringeria, 18, 201–245.
Arendt, Y. A. (1974). Morskie lilii. Tsirtokrinidy. Akad. Nauk SSSR, Paleont. Inst., Trudy, 144, 251 pp. Izdatelstvo “Nauka” (Moskva). [Sea lilies, Cyrtocrinids].
Ausich, W. I., Donovan, S. K., Hess H., Simms, M. J. (1999). In H. Hess, W. I. Ausich, C. E. Brett & M. J. Simms (Eds.). Fossil crinoids (pp. 41–49). Cambridge University Press: Cambridge.
Ausich, W. I., & Simms, M. J. (1999). Ecology and Ecological Interactions, In H. Hess, W. I. Ausich, C. E. Brett & M. J. Simms (Eds.). Fossil crinoids (pp. 55–59). Cambridge University Press: Cambridge.
Bigot, A. P. D. (1938). Crinoïdes du Bathonien du Calvados. Annales de Paléontologie, 27, 1–38.
Boule, M. (1910–1911). Types du Prodrome (Suite). Annales de Paléontologie, 5, 93–116 (1910); 6, pl. 22 (1911).
Bourseau, J.-P., Améziane-Cominardi, N., Avocat R., & Roux, M. (1991). Echinodermata: Les Crinoïdes pédonculés de Nouvelle-Calédonie. In A. Crosnier (Ed.). Résultats des Campagnes MUSORSTOM 8. Mémoires du Muséum national d’Histoire naturelle, (A) 151, 229–333, Paris.
Bromley, R. G. (1981). Concepts in ichnotaxonomy illustrated by small round holes in shells. Acta Geológica Hispánica, 16, 55–64.
Charbonnier, S. (2009). Le Lagerstätte de la Voulte, un environnement bathyal au Jurassique. Mémoires du Muséum national d’Histoire naturelle 199. 272 pp. Paris.
Charbonnier, S., Vannier, J., Gaillard, C., Bourseau, J.-P., & Hantzpergue, P. (2007). The La Voulte Lagerstätte (Callovian): evidence for a deep water setting from sponge and crinoid communities. Palaeogeogr Palaeoclimatol Palaeoecol, 250, 216–236.
d’Orbigny, A (1850–1852). Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés faisant suite au cours élémentaire de paléontologie et de géologie stratigraphique, 1 (1849 [1850]), 394 pp.; 2 (1850[1852]), 427 pp.; 3 (1852), 196 pp. + table alphabétique et synonymique des genres et des espèces, 1–189. Masson: Paris.
de Loriol, P. (1877–1879). Monographie des crinoïdes fossiles de la Suisse. Mémoires de la Société Paléontologique Suisse, 4, 1–52; 5, 3–124; 6, 125–300.
de Loriol, P. (1882–1889). Paléontologie française, Série. 1, Animaux invertébrés. Terrain jurassique, 11, Crinoïdes, 1 (1882–1884) 627 pp.; 2 (1884–1889) 580 pp. G. Masson: Paris.
Donovan, S. K., Widdison, R. E., Lewis, D. N., & Fearnhead F. E. (2010). The British silurian crinoidea. Part 2. (pp. 47–133) Monograph of the Palaeontological Society: London. Addendum to Part 1 and Cladida. Monograph of the Palaeontological Society, London 47–133 (Publ.635, part of Vol. 164 for 2010).
Dumortier, E. (1871). Sur quelques gisements de l’Oxfordien inférieur de l’Ardèche. 84 pp. F. Savy, éditeur: Paris; Josserand Librairie: Lyon.
Elmi, S. (1967). Le Lias supérieur et le Jurassique moyen de l’Ardèche. Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon 19 (fasc. 1) 1–256, (fasc. 2) 257–507.
Etter, W. (2002). La Voulte-sur-Rhône: Exquisite Cephalopod Preservation. In D. J. Bottjer, W. Etter, J. W. Hagadorn, & C. M. Tang (Eds.), Exceptional fossil preservation (pp. 293–305). New York: Columbia University Press.
Gislén, T. (1924). Echinoderm studies. Zoologisk Bidrag från Uppsala, 9, 330 pp.
Goldfuss, G. A. (1826–1833). Petrefacta Germaniae. Abbildungen und Beschreibungen der Petrefacten Deutschlands und der Angränzenden Länder, unter Mitwirkung des Herrn Grafen Georg zu Münster, herausgegeben von August Goldfuss. Band 1 (1826–18833), Divisio prima. Zoophytorum reliquiae, 1–114; Divisio secunda. Radiariorum reliquiae, 115–221 [Echinodermata]. Arnz & Co.: Düsseldorf.
Helm, C., Reuter, M. & Schülke, I. (2003). Comatulid crinoids from Late Jurassic reefal deposits of NW Germany (Lower Saxony Basin). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 2003, 513–528.
Hess, H. (1960). Neubeschreibung von Geocoma elegans (Ophiuroidea)aus dem unteren Callovien von La Voulte-sur-Rhône (Ardèche). Eclogae Geol Helv, 53, 335–385.
Hess, H. (1975). Die fossilen Echinodermen des Schweizer Juras. Veröffentlichungen aus dem Naturhistorischen Museum Base,l 8, 130 pp.
Hess, H. (1999). Middle Jurassic of northern Switzerland. In H. Hess, W. I. Ausich, C. E. Brett, & M. J. Simms (Eds.), Fossil crinoids (pp. 203–215). Cambridge: Cambridge University Press.
Hess, H. (2004). Sclerocrinus Jaekel, 1891 (Crinoidea, Cyrtocrinida): proposed precedence over Gammarocrinites Quenstedt, 1857. Bulletin of Zoological Nomenclature, 61(4), December 2004, 251–253.
Hess, H. (2006). Crinoids from the Lower Jurassic (Pliensbachian) of Arzo, southern Switzerland. Schweizerische Paläontologische Abhandlungen, 126, 143 pp.
Hess, H. (2008). Cyclocrinus, an enigmatic Jurassic–Cretaceous crinoid. Swiss J Geosci, 101, 465–481.
Hess, H., & Gale, A. S. (2010). Crinoids from the Shenley Limestone (Albian) of Leighton Buzzard (Bedfordshire, UK). J Syst Paleontol, 8, 427–447.
Hess, H. & Messing, C. G. (2011). Treatise on invertebrate paleontology, Part T. In: W. I. Ausich (Ed.) Echinodermata 2, revised, Crinoidea, vol. 3 (261 pp). The University of Kansas Paleontological Institute: Lawrence.
Hess, H., & Spichiger, P. U. (2001). Argoviacrinus rarissimus n. g. n. sp., a new crinoid (Echinodermata) from the Middle Oxfordian of northern Switzerland. Eclogae Geol Helv, 94, 489–494.
Jaekel, O. (1891). Ueber Holopocriniden mit besonderer Berücksichtigung der Stramberger Formen. Zeitschrift der Deutschen geologischen Gesellschaft, 43, 557–670.
Jäger, M. (1993). Die Crinoiden aus dem Pliensbachium vom Rhein-Main-Donau-Kanal bei Sulzkirchen. Teil 2: Gestielte Cyrtocrinina und Millericrinina: Amaltheocrinus, Shroshaecrinus? Plicatocrinus und Nachtrag zu Teil 1 (Eudesicrinus). Archaeopteryx, 11, 73–104. Eichstätt.
Jäger, M. (1995). Echinodermata aus dem Ober-Toarcium und Aalenium Deutschlands. I. Crinoidea: Cyrtocrinina und Millericrinina. Stuttgarter Beiträge für Naturkunde, (B) 226, 51 pp.
Klikushin, V. G. (1987). Crinoids from the Middle Liassic Rosso ammonitico beds. Neues Jb Geol Paläontol Abh, 175, 235–260.
Klikushin, V. G. (1996). Late Jurassic Crinoids from Sudak environs (Crimea). Palaeontographica, Abteilung A, 238, 97–151.
Kristan-Tollmann, E. (1988). Palaeocomaster styriacus n. sp., eine Schwebcrinoide (Comatulida) aus dem alpinen Lias des Salzkammergutes, Österreich. Geol Palaeontol, 22, 81–87.
Lissajous, M. (1912). Jurassique mâconnais, Fossiles caractéristiques. Bulletin trimestriel de la Société d’histoire naturelle de Mâcon, 3 (3–16), 208 pp.
Manni, R., Nicosia, U., & Riou, B. (1985). Rhodanometra lorioli n. gen. n. sp. and other Callovian crinoids from La Voulte-sur-Rhône (Ardèche, France). Geol Romana, 24, 87–100.
Nicosia, U. (1991). Mesozoic crinoids from the north-western Turkey. Geol Romana, 27, 389–436.
Oppel, A. (1865). Geognostische Studien in dem Ardèche Departement. Paläontologische Mittheilungen aus dem Museum des Königlich Bayerischen Staates, 1, 305–322.
Quenstedt, F. A. (1876). Petrefactenkunde Deutschlands. 1. Abt., 4. Band, Echinodermen (Asteriden und Encriniden), viii + 742 pp. Fues’s Verlag: Leipzig.
Radwanska, U. (2007). A rare comatulid crinoid, Semiometra petitclerci (Caillet, 1923) from the Upper Oxfordian of Poland. Acta Geologica Polonica, 57, 161–167.
Rasmussen, H. W. (1978). Articulata. In R. C. Moore & C. Teichert, (Eds.) Treatise on invertebrate paleontology. Pt. T, Echinodermata 2, vol. 3, T813-T928. Geological Society of America and University of Kansas Press: Boulder.
Roman, F. (1950). Le Bas-Vivarais.Géologie régionale de la France, VI. 150 pp. Hermann & Cie: Paris.
Sayn, G., & Roman, F. (1928). Monographie stratigraphique et paléontologique du Jurassique moyen de la Voulte-sur-Rhône. Travaux du Laboratoire de Géologie de la Faculté des Sciences de Lyon II, 13, 1–165.
Simms, M. J. (1989). British Lower Jurassic crinoids, vol. 581 (103 pp.). Monograph of the Palaeontographical Society: London.
Valette, A. (1927). Note sur quelques débris de Crinoïdes fossiles de la Catalogne. Bulletin de la Institucio Catalana d’Historia Natural, 7, 24–27.
Ziegler, B. (1977). The “White” (Upper) Jurassic in Southern Germany. Stuttgarter Beiträge zur Naturkunde, (B) 26, 79 pp.
Acknowledgments
I am greatly indebted to Andy Gale for providing the bulk samples and information on the sites, including sketches of the beds sampled. I also thank Walter Etter for the line drawings and useful discussions. E. Robert, Grenoble University, kindly provided specimens for study. S. K. Donovan helped with information on epizoans. The SEM photographs were by E. Bieler, Zentrum für Mikroskopie of the University of Basel. I would like to express my sincere thanks to S. K. Donovan and an unknown reviewer for their thorough reviews and constructive comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hess, H. Crinoids from the Middle Jurassic (Bajocian–Lower Callovian) of Ardèche, France. Swiss J Palaeontol 131, 211–253 (2012). https://doi.org/10.1007/s13358-012-0044-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-012-0044-9