Special issue: Recent advances in Cambrian to modern cephalopod research I
- Published:
Ontogeny and morphology of the retrochoanitic septal necks of Agoniatites vanuxemi from the Middle Devonian Cherry Valley Limestone
Swiss Journal of Palaeontology volume 134, pages 233–244 (2015)
Abstract
Retrochoanitic septal necks represent the primitive condition in the Ammonoidea. In this study, we examine the morphology of the septal siphuncular complex of specimens of Agoniatites vanuxemi (Hall 1879) from the Cherry Valley Limestone of New York State, USA. Specimens were embedded in epoxy, ground to a median plane and etched in 5 % HCl. The phosphatized membranes allow us to study the formation of the septal necks. Septal–siphuncular membranes occur next to the septal neck. They are adapical of the septum at the deepest point of the adorally projected ventrolateral saddle. Similar membranes occur on the adapical side of the septum on the dorsum in the deepest part of the adorally projected septa. The septal neck morphology on both the dorsal and ventral sides is well preserved. The septal neck begins in the same position on its dorsal and ventral sides, but projects farther adorally on the ventral side. By the third whorl, the adoral projection has doubled in size and is one-fourth to one-third the length of the chamber. This may be related to the deepening of the ventrolateral saddle through ontogeny. Auxiliary deposits are also present on the inside of the septal necks. They occur inside the dorsal and ventral sides of the septal neck at the adapical end. On some septa they appear to extend adapically beyond the end of the septal neck. A model for the morphogenesis of retrochoanitic septal necks is proposed based on the morphology observed in these specimens and previously published models.
Introduction
Retrochoanitic septal necks appear in the phylogenetically oldest members of the Ammonoidea (Miller et al. 1957), as well as in their orthocerid ancestors (Teichert 1964; Becker and Kullmann 1996). It is the only form found in Devonian ammonoids (including the Bactritida, Agoniatitina, Anarcestina, and Tornoceratina) (Branco 1879–1880; Drushchits et al. 1976). Tanabe and Landman (1996) surveyed available specimens and literature on septal neck morphology. Their discussion on Devonian taxa is less than one-half page, mainly because of lack of research on these forms. (See Tanabe and Landman 1996, p. 138 for literature used in their study.) The models they created for retrochoanitic septal necks are largely based on the better preserved and more intensively studied Goniatitina. In fact, it is only when retrochoanitic septal necks are associated with membranes (as in prolecanitids and goniatites) (Landman et al. 2006; Polizzotto et al. 2007) or ontogenetically change into another form (as in goniatites) (Miller and Unklesbay 1943; Doguzhaeva and Mutvei 1986) do they garner more attention. In this paper, we examine the early ontogeny of retrochoanitic septal necks in the Middle Devonian ammonoid Agoniatites vanuxemi (Hall, 1879). It is hoped that this study will promote further investigation of these phylogenetically older taxa, and offer greater understanding of how these structures formed and evolved.
The ammonoids used in this study are from the Middle Devonian Cherry Valley Limestone (New York State, USA). The formation is exposed within a band of Middle Devonian rocks that stretch across the length of central New York State (Fig. 1). The Cherry Valley Limestone is the topmost member in the lower part of the Marcellus Formation (Rickard 1952; Griffing and Ver Straeten 1991). It is described as a dark, argillaceous, massive, nodular, styliolinid-cephalopod packstone, containing a mostly pelagic fauna. A. vanuxemi is the most abundant cephalopod present, with numbers estimated to be in the billions across New York State (Anderson et al. 1988). Based on the large number of specimens occurring in single beds, in contrast to the rest of the Cherry Valley Limestone, most workers have suggested a mass mortality event (Anderson et al. 1988; Van Dyk 1988; Whiteley and Kloc 1999; Klofak 2002). In other parts of the Cherry Valley Limestone, specimens occur isolated in the rock with a variety of orientations, though most lie horizontally (Cottrell 1973, unpublished Masters Thesis). The orientation is thought to reflect a variety of factors including the condition of the shell, amount of air remaining in the chambers, current velocity, water depth, nature of the substrate, and bottom topography (for further discussion of ammonoid taphonomy, see Reyment 1970; Raup 1973; Crick 1983; Wani and Ikeda 2006; Olivero 2007; De Baets et al. 2008).

Map of New York State, USA. The dark band represents exposures of Middle Devonian rocks. The arrow points to the location of the Seneca Stone Quarry
Materials and methods
The specimens in this study were collected in the Seneca Stone Quarry. Two specimens were previously studied and occur in the Lagerstätten bed (AMNH 53356 and AMNH 53357, Klofak and Landman 2010). The remaining specimens were removed from pieces of bedrock collected in the quarry. They are all phragmocones; no body chambers are preserved. PRI 68137 shows the type of preservation common in these specimens (Fig. 2). The specimen consists of five whorls, the outermost of which is mud filled. The fourth whorl is hollow. The septa are missing and the void is lined with large calcite crystals. The inner three whorls are surrounded by hollow chambers. PRI 68139, though not as complete, is similar to PRI 68137 in preservation. PRI 68138 is different in its preservation; it is smaller, with four whorls. Only the last one-quarter whorl is exposed. It appears to be almost completely mud filled. The hollow spaces, found in some specimens, are believed to be the result of a vertical settling position and gases trapped in the chambers (Cottrell 1973). These hollow chambers allowed for easy extraction of the inner whorls used in this study.

Agoniatites vanuxemi. PRI 68137. Specimen as it occurs in limestone block. Arrow indicates adoral direction. V void space where septa are missing, E early whorls removed for study. Scale bar 1 cm
Once the inner whorls were removed from the specimens, they were embedded in epoxy and ground to a median plane, or in some cases just above the median plane. They were then polished and etched with 5 % HCl for approximately 10–15 s. The specimens were then examined using a Zeiss EVO-60-EP-SEM. Energy Dispersive X-ray Spectrometry (EDS) was used to determine elemental composition. Specimens that were examined above the median plane were re-polished and re-etched and again studied with the SEM. For some specimens, this process was repeated several times until the median plane was achieved.
For this study, the count of the number of whorls includes the ammonitella, i.e., the first whorl is the ammonitella, the second whorl is the first post-embryonic whorl and so forth. Specimens are reposited in the American Museum of Natural History, New York (AMNH) and Paleontological Research Institute, Ithaca, New York (PRI).
Results
The inner whorls of the specimens are generally calcite filled. In some of the chambers the calcite fill is not complete, leaving some void space. The ammonitellas are not well preserved. In PRI 68137, only traces remain and the area is filled with calcite. In PRI 68138, the initial chamber is filled with pyrite and calcium phosphate. Pyrite, dolomite, and silica are also present in both the mud and the calcite fill. No original aragonite is preserved. The shell and septa are replaced with blocky calcite. The organic linings of the shell and septa, as well as organic structures such as the siphuncle and membranes are preserved as calcium phosphate. The phosphatized organic membranes allow us to study the formation of the septal necks and associated structures (Klofak and Landman 2010).
Septal–siphuncular membranes (for the original definition of these membranes see Landman et al. 2006) were observed in three specimens (PRI 68137, PRI 68138, AMMH 53357); they are always above the median plane; in other words, to the side of the siphuncle. They occur in the deepest part of the ventrolateral saddle (Fig. 3). The best examples are visible in the second whorl of PRI 68138. They attach to the adapical side of the septa and stretch to the ventral shell wall (Fig. 3c, d). They are about 50 μm in length, equivalent to ~0.04 times the height of the chamber. The same membranes appear in PRI 68137 (Fig. 3a) and AMNH 53357 (Fig. 3b), both in the second whorl. In a previous study, these membranes are associated with septa in the ammonitella (see Klofak and Landman 2010, fig 9g, h). Septal–siphuncular membranes may also occur in the ammonitella of PRI 68138, but preservation is too poor to be sure.

Agoniatites vanuxemi. Septal–siphuncular membranes. a PRI 68137, second whorl. Scale bar 20 μm. b AMNH 53357, second whorl. Scale bar 20 μm. c, d PRI 68138, second whorl. c Scale bar 100 μm. d Scale bar 100 μm. SSM septal–siphuncular membranes, V ventral surface of chamber, S septum, arrow adoral direction
Similar looking types of septal membranes occur on the dorsal part of the chamber. They have been observed in several specimens (Fig. 4). As with the septal–siphuncular membranes described above, these dorsal-septal membranes, as defined here, occur above the median plane of the sectioned specimen. They are located on the adapical side of the septum and lie deep in the forward projected part of the septum, near the dorsal shell wall. Two parallel membranes are usually present. They stretch from the adapical side of the septum to the dorsal shell wall. Figure 4c, e, f shows examples of these membranes on two successive septa in the second whorl of PRI 68138. The longer of these membranes measures 60 μm. These same dorsal-siphuncular membranes are observed on three septa, exactly one whorl later in the third whorl (Fig. 4d).

Agoniatites vanuxemi. Dorsal-septal membranes. a AMNH 53357, second whorl. Scale bar 20 μm. b PRI 68137, dorsal-septal membranes in the second whorl. Scale bar 100 μm. c–f PRI 68138. c Dorsal-septal membranes on three septa in the third whorl. Scale bar 100 μm. d Dorsal-septal membranes in second whorl. Scale bar 100 μm. e Close-up of septal membrane on left, from c. Scale bar 10 μm. f Close-up of septal membranes on right, from c. Scale bar 10 μm. DSM dorsal-septal membranes, D dorsal surface of chamber, V ventral surface of preceding whorl, S septum, * same place on specimen in c and d. Arrow adoral direction
Septa and septal neck morphology are also preserved in these specimens. Their morphology is outlined by the phosphatized organic membranes, which line the septa. These membranes reveal excellent details about the structure of both the dorsal and ventral sides of the septal necks. Figure 5 shows septal necks from the second whorl of three specimens. Viewed in median cross section, the adapical end of the septal necks on both the dorsal and ventral sides begins at a point equally distant from the bend on the septal neck. The adapical end of the septal neck is pointed and is usually covered on the inside by a calcium phosphate deposit (Fig. 5d–f). This deposit can sometimes extend adapically beyond and around the outside of the beginning point of the septal neck (Fig. 5a, b). The ventral side of the septal neck is harder to trace as it lies against and sometimes seems to merge into the ventral wall of the specimen (Fig. 5b, d). A discernible gap is visible at the adapical end. Adorally, the septal neck moves closer to the ventral shell wall, thinning and merging into the wall. A calcium phosphate deposit occurs on the adapical end on the inside of the neck, opposite the deposit on the dorsal side of the neck. In a specimen where several septal necks are sectioned just above the median plane, details of the microstructure of the septal necks are reflected in the preserved phosphatized linings. At the adoral end, the inner surface of the lining appears smooth, reflecting the smooth regular shell surface of the septum. At the adapical end the texture is rough, reflecting an area where the tissue is less homogeneous, possibly mixed with organic material, now preserved as calcium phosphate deposits (Fig. 6a, b). The same textural variation is visible in phosphatized membranes of septal necks sectioned just below the median plane (Fig. 6c, d).

Agoniatites vanuxemi. Septal necks. a, b PRI 68137. a Septal neck in second whorl. Dorsal side of septal neck is well preserved. Auxiliary deposits on the dorsal and ventral sides of neck extend adapically beyond the end of the septal neck. Septum split by calcite seam below dorsal side of septal neck. Scale bar 200 μm. b Another septal neck in the second whorl. Auxiliary deposits are well preserved on dorsal and ventral sides. Scale bar 100 μm. c AMNH 53356. Septal neck with organic membranes extending across siphuncle. Scale bar 100 μm. d–f PRI 68138. d Well-preserved septal neck from second whorl. Scale bar 30 μm. e Close-up from d. Ventral side of septal neck with auxiliary deposit preserved. Scale bar 10 μm. f Dorsal side of septal neck with well-preserved auxiliary deposit. Scale bar 10 μm. S septum, V venter, DSN dorsal part of septal neck, VSN ventral part of septal neck, aux auxiliary deposits, arrow adoral direction

Agoniatites vanuxemi. PRI 68139. Septal necks cut above/below the median plane. a Rough texture visible at the adapical end of septal neck; smooth texture at adoral end. Scale bar 100 μm. b Rough texture visible at the adapical end of septal neck. Scale bar 100 μm. c, d Section below median plane. Rough texture visible at adapical end; smooth texture of septal neck visible at adoral end. Scale bar 200 μm. d Close-up from c, showing rough texture at adapical end of septal neck. Scale bar 20 μm. SN septal neck, rt rough texture, st smooth texture, V venter, S septum, arrow adoral direction
PRI 68139 is a unique specimen. It is split through the center by a large dolomite vein, but the whorls on either side of the vein are preserved and can be followed. There are fewer membranes, but the ones that are present preserve an important septal morphology. The septal necks described in all the studied specimens appear in the second whorl. In PRI 68139, the morphology of the septal neck in the second whorl is the same as that in all of the other studied specimens (Fig. 7a). This can be contrasted with the septal necks in the third whorl of PRI 68139, where the ventral sides of the septal necks are much longer than on the dorsal sides (Fig. 7b–d). Like all of the preceding septal necks, the adapical ends of the dorsal and ventral sides of the septal necks begin equidistant from the bend in the septal necks. However, the ventral side of the septal necks extents adorally into the next chamber for one-fourth to one-third of the chamber length (Fig. 7b). This adoral projection is approximately one-half the length of the ventral septal neck. Most of the projection lies against the ventral shell wall, while the adapical end of the septal neck is away from the shell wall (Fig. 7c, d). (Chamber length is measured on the dorsal side of the septal neck from the adapical tip of the septal neck of one septum to the adoral bend of the septal neck of the preceding septum.)

Agoniatites vanuxemi. PRI 68139. a Septal neck in the second whorl. Scale bar 100 μm. b–d Septal necks in the third whorl. b Two septal necks. The length of the dorsal extension of the septal neck into the chamber is visible in this view. Scale bar 20 μm. c Close-up of septal neck in b (1). Adoral extent of dorsal side of septal neck is marked by a star. Scale bar 200 μm. d Close-up of septal neck in b (2). Adoral extent of dorsal side of septal neck is marked by a star. Auxiliary deposits are preserved. Scale bar 200 μm. S septum, V venter, DSN dorsal part of septal neck, VSN ventral part of septal neck, aux auxiliary deposits, arrow adoral direction
Discussion
The septal necks of A. vanuxemi have many similarities to those in other ammonoid species with retrochoanitic septal necks, for at least part of their ontogeny. For example, long septal necks have been documented in Agoniatites, Anarcestes, Latanarcestes (Spath, 1933) Prouddenites, Uddenites (Miller and Unklesbay, 1943), and Agathiceras (Miller and Unklesbay, 1943; Doguzhaeva and Mutvei, 1986).
And while no shell microstructure is preserved, we can infer that the first and last deposited layer of the septum is organic. The phosphatized organic layers provide information about the morphology of the dorsal and ventral sides of the septal necks. In median cross section the two sides are not the same length. They begin adapically the same distance from the bend of the septal neck, but the ventral side projects adorally into the next chamber (Fig. 8). The distance of the adoral projection doubles within one whorl (Fig. 9). This is likely a function of the shape of the septa. As the ventrolateral saddle deepens, the ventral side of the septal neck lengthens.

Diagrammatic cross-sectional representation of the septal necks of Agoniatites vanuxemi. S siphuncular tube, VSN ventral septal neck, DSN dorsal septal neck, aux auxiliary deposits, VW ventral shell wall, arrow indicates the adoral direction

Schematic representations of the septal necks contrasting the length of the dorsal and ventral sides of the septal necks through ontogeny. a Septal neck from the second whorl. The dorsal septal neck is only a little longer than the ventral septal neck. b In a later whorl the ventral side of the septal neck is much longer and extends into the next chamber. S siphuncle, VSN ventral septal neck, DSN dorsal septal neck, VW ventral shell wall, arrow indicates the adoral direction
There is also evidence of auxiliary deposits, but unlike those previously described by Miller and Unklesbay (1943), Kulicki (1979), and Tanabe and Landman (1996), the ones in A. vanuxemi seem to be largely made of originally organic material (Kulicki and Mutvei 1982). They are positioned on the adapical end of the inside of the septal neck, and in some cases extend adapically beyond the end of the septal neck (Figs. 5a, b, 8). Reconstructions of these deposits vary based on the taxa and how the authors choose to reconstruct them (Tanabe and Landman 1996; Doguzhaeva and Mutvei 1986). For example, Doguzhaeva and Mutvei (1986, Fig. 1A) reconstructed a retrochoanitic septal neck of a ceratite, with the auxiliary deposit at the adoral end of the septal neck with a separate attachment site for the connecting ring at the adapical end. The reconstruction by Tanabe and Landman (1996, fig. 2a) is more similar to what we observed in A. vanuxemi. The auxiliary deposits occur closer to the adapical end of the septal neck. In A. vanuxemi, the auxiliary deposits also extend beyond the adapical tip of the septal neck. The connecting ring may attach to the septal neck at a single site, the auxiliary deposit, and pass through the septal neck (Miller and Unklesbay 1943). There is no evidence for a point of attachment of an organic structure at the adoral end of the septal neck, based on the observed texture of the septal necks (Fig. 6). If a gap exists between the adoral and adapical points of attachment of the connecting ring it is, at most, the length of the auxiliary deposit (approximately 50 μm).
Membranes play an important role in the formation of the septa in Agoniatites (Fig. 10). The only reason the morphogenesis of the septa can be studied is because of the presence of organic linings on the dorsal and ventral surfaces of the septa and shell. In addition to these membranes, septal membranes also occur. They are associated with septa in both the embryonic (Klofak and Landman, 2010) and post-embryonic stages. The septal–siphuncular membranes appear next to the siphuncle, on the adapical side of the septum in the deepest part of the forward projection of the ventrolateral saddle (Fig. 10b). The dorsal-septal membranes are similar in form to the septal–siphuncular membranes and appear adapical of the septum in the forward projection on the dorsal side of the septum. The septal–siphuncular membranes resemble those described by Landman et al. (2006, figs. 8d, 13c) in the prolecanitid Akmilleria electraensis (Plummer and Scott, 1937), but in A. electraensis the membranes appear on the adoral side of the septa in a deep backward projecting part of the septum, next to the siphuncle. Landman et al. (2006) proposed a number of possible functions for these membranes. And while none can be completely ruled out, the use of these membranes for facilitating liquid removal seems the most reasonable (see also Kulicki 1979; Weitschat and Bandel 1991; Checa 1996). The efficiency of draining the liquid from the deep groove formed behind the septum may have been enhanced by the presence of these membranes. They may also have formed as a function of the forward movement of the animal as it grew in the shell. The back part of the mantle may have retained some flexibility and may not have maintained the deep pointed adoral flexure where it attached to the shell. When it reached the position for the next septum to form, the margins may have bent forward in small increments until the septum reached the desired shape. This type of movement may have aided in the mantle’s ability to increase the depth of the ventrolateral saddle, as well as deepen the convexity of the lateral lobe.

Three-dimensional view of septal necks in Agoniatites vanuxemi. a View of septal neck. b Close-up of ventral side of septal neck showing the placement of the septal–siphuncular membranes. S septum, SN septal neck, aux auxiliary deposits, VW ventral shell wall, ST siphuncular tube, SSM septal–siphuncular membranes. Arrow adoral direction. Modified from Tanabe and Landman (1996, fig. 1)
Based on the data collected from A. vanuxemi, and following the lead of Tanabe and Landman (1996, Fig. 11), we propose our own morphogenetic model for septal formation. In creating the model we have relied on our observations and the observations of others (Mutvei 1964; Tanabe et al. 1982; Tanabe and Landman 1996). Klug et al. (2008) further modified septal morphogenesis models to include the function of mantle muscles in the translocation of the animal in the shell. The presence of these muscles was based on detailed analysis of both modern Nautilus (tissue and shell) and impressions left on fossils of nautiloids and early ammonoids. And while muscle attachment and detachment plays a role in translocation, the phosphatized organic linings and membranes preserved in our specimens offer no clues as to their role. Our model is more focused on the mechanism used in the production of the organic membranes.
-
1.
The rear mantle secretes its final organic layer, detaches from the adoral face of the septum and septal neck, and moves forward (Fig. 11a).
Fig. 11 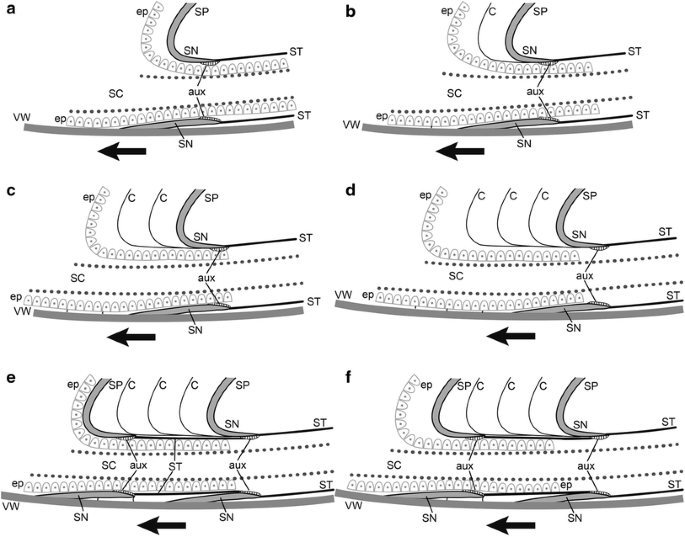
Morphogenetic model showing successive development of the retrochoanitic septal neck of A. vanuxemi (a–f). See text for details. C cameral membranes, ep septal epithelium, SC siphuncular cord, ST siphuncular tube, aux auxiliary deposits, SP septum, SN septal neck, VW ventral shell wall, arrow indicates the adoral direction. Modified from Tanabe and Landman (1996, fig. 11A)
-
2.
As the animal moves forward, the rear mantle pauses and produces an organic membrane. After the membrane is secreted, the animal moves forward again (Fig. 11b).
-
3.
The animal continues to move forward, pausing periodically and producing membranes (Fig. 11c).
-
4.
When the animal moves forward the appropriate distance, it stops its forward movement, except near the shell wall where it continues to move forward in very small increments, producing the septal–siphuncular and dorsal-septal membranes. Once the septal shape is attained, the dorsal layer is produced on the adapical side. The siphuncular epithelium secretes the rest of the siphuncular tube and the auxiliary deposits on the adoral side of the already formed septum (Fig. 11d).
-
5.
A new septum is secreted, with an organic membrane on its adoral side (Fig. 11e).
-
6.
The rear mantle detaches from the adoral face of the septum and septal neck and the process begins again (Fig. 11f).

Conclusions
With no shell microstructure preserved, alternate means must be found to study the morphology and ontogeny of the septa in Devonian ammonoids, including A. vanuxemi. One method is to use the organic membranes that line the shell wall and septa of the specimens. The presence of these membranes is well established in many taxa (Grandjean 1910; Shoulga-Nesterenko 1926; Shindewolf 1968; Erben and Reid 1971; Bayer 1977; Weitschat and Bandel 1991; Tanabe et al. 1993; Tanabe and Landman 1996; Landman et al. 2006; Polizzotto et al. 2007) and with this study, they can be extended to the Agoniatitaceae. The shape of the septal necks of A. vanuxemi is documented and changes in its morphology (particularly on the ventral side) are visible. These changes would seem to be associated with changes in the shape of the septa (the deepening of the ventrolateral saddle). The presence of septal–siphuncular membranes and dorsal-septal membranes might also, in part, be a reflection of the forward movement of the animal during the process of septal formation.
With these data, it is possible to compare the morphology of retrochoanitic septal necks in one of the oldest taxa with retrochoanitic septal necks in younger taxa, many of which do not retain retrochoanitic necks throughout ontogeny. A review of the literature does not reveal the increase in length of the ventral side of the septal neck in younger taxa. Most of these studies focus on the changes that occur on the dorsal side of the septal neck. This is due, in part, to preservational bias. Tanabe et al. (1993) noted that septal necks were initially classified based only on the direction in which the dorsal side projects. Generally, unless the ventral portion of the septal neck lies away from the shell wall, it is often impossible to separate this structure from the shell wall, or the structure is lost entirely (Kulicki 1979; Doguzhaeva and Mutvei 1986). The data in this study suggest that these changes may be important and need further clarification. Such investigations need to incorporate data on the shape and changes in septal morphology, especially in the area near the siphuncle.
References
Anderson, E. J., Brett, C. E., Fisher, D. W., Goodwin, P. W., Kloc, G. J., Landing, E., & Lindemann, R. H. (1988). Upper Silurian to Middle Devonian stratigraphy and depositional controls, East-Central New York. In E. Landing (Ed.), The Canadian paleontology and biostratigraphy seminar, New York State Museum Bulletin, 462, 111–134.
Bayer, U. (1977). Cephalopoden-Septen. I. Konstruktionsmorphologie des Ammoniten-Septums. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 154, 290–366.
Becker, R. T., & Kullmann, J. (1996). Paleozoic ammonoids in space and time. In N. H. Landman, K. Tanabe, & R. A. Davis (Eds.), Ammonoid paleobiology (pp. 711–753). New York: Plenum Press.
Branco, W. (1879–1880). Beitraege zur Entwickelungsgeschichte der fossilen Cephalopoden. Palaeontographica, 26, 15–50; 27, 17–81.
Checa, A. (1996). Origin of intracameral sheets in ammonoids. Lethaia, 29, 61–75.
Cottrell, J. F. (1973). The paleoecological significance of inclined cephalopods, Cherry Valley Limestone (Devonian), New York. Unpublished Master’s Thesis, University of Rochester, pp. 23, 7pls.
Crick, R. E. (1983). The practicality of vertical cephalopod shells as paleobathymetric markers. Geological Society of America Bulletin, 94, 1109–1116.
De Baets, K., Cecca, F., Guiomar, M., & Verniers, J. (2008). Ammonites from the latest Aalenian–earliest Bathonian of La Baume (Castellane area, SE France): Palaeontology and biostratigraphy. Swiss Journal of Geoscience, 101, 563–578.
Doguzhaeva, L., & Mutvei, H. (1986). Retro- and prochoanitic septal necks in ammonoids, and transition between them. Palaeontographica Abteilung A, 195(1–3), 1–18.
Drushchits, V. V., Bogoslovskaya, M. F., & Doguzhayeva, L. A. (1976). Evolution of septal necks in the Ammonoidea. Paleontological Journal, 1, 41–56.
Erben, H. K., & Reid, R. E. H. (1971). Ultrastructure of shell, origin on conellae, and siphuncular membranes in an ammonite. Biomineralization Research Reports, 3, 22–31.
Grandjean, F. (1910). Le siphon des ammonites et des belemnites. Bulletin de la societé Géologique de France, 10, 496–519.
Griffing, D. H., & Ver Straeten, C. A. (1991). Stratigraphy and depositional environments of the lower part of the Marcellus Formation (Middle Devonian) on eastern New York State. New York State Geological Association 63rd annual meeting, SUNY Oneonta, 205–234.
Hall, J. (1879). Descriptions of Devonian fossils; Gasteropoda, Pteropoda, Cephalopoda of the Upper Helderberg, Hamilton, Portage and Chemung groups (Vol. 5, pt. 2, pp. I–xv, 1–492, pls. 1–113). New York Geological Survey, Palaeontology of New York.
Klofak, S. M. (2002). Size classes in a Devonian ammonoid from the Middle Devonian Cherry Valley Limestone of New York State, U.S.A. In H. Summesberger, K. Histon & A. Daurer (Eds.), Cephalopods—Present and past. Abhandlungen der Geologischen Bundesanstalt, 57, 443–457.
Klofak, S. M., & Landman, N. H. (2010). Some exceptionally well preserved specimens of Agoniatites vanuxemi from the Middle Devonian Cherry Valley Limestone of New York State, U. S. A. In K. Tanabe, Y. Shigeta, T. Sasaki & H. Hirano (Eds.), Cephalopods—Present and past (pp. 93–103). Tokai University Press.
Klug, C., Meyer, E. P., Richter, U., & Korn, D. (2008). Soft-tissue imprints in fossil and recent cephalopod septa and septum formation. Lethaia, 41, 477–492.
Kulicki, C. (1979). The ammonite shell: Its structure, development and biological significance. Palaeontologia Polonica, 39, 96–141, pls. 24–48.
Kulicki, C., & Mutvei, H. (1982). Ultrastructure of the siphonal tube in Quenstedtoceras (Ammonitina). Stockholm Contributions in Geology, 37, 129–138.
Landman, N. H., Polizzotto, K., Mapes, R. H., & Tanabe, K. (2006). Cameral membranes in prolecanitid and goniatitid ammonoids from the Permian Arcturus Formation, Nevada, USA. Lethaia, 39, 365–379.
Miller, A. K., Furnish, W. M., & Schindewolf, O. H. (1957). Paleozoic Ammonoidea. In R. C. Moore (Ed.), Treatise on Invertebrate Paleontology, Part L, Mollusca 4 (pp. L11–L79). Lawrence, KS: Geological Society of America and University of Kansas Press.
Miller, A. K., & Unklesbay, A. G. (1943). The siphuncle of Late Paleozoic ammonoids. Journal of Paleontology, 17, 1–25.
Mutvei, H. (1964). On the shells of Nautilus and Spirula with notes on the shell secretion in non-cephalopod molluscs. Arkiv for Zoologi, 16(14), 221–278.
Olivero, E. B. (2007). Taphonomy of ammonites from the Santonian-Lower Campanian Santa Marta Formation, Antartica: Sedimentological controls on vertically embedded ammonites. Palaios, 22, 586–597.
Plummer, F. B., & Scott, G. (1937). Upper Paleozoic ammonites in Texas. The geology of Texas. The University of Texas Bulletin, 3701, 1–516.
Polizzotto, K., Landman, N. H., & Mapes, R. H. (2007). Cameral membranes in Carboniferous and Permian goniatites: Description and relationship to pseudosutures. In N. H. Landman, R. H. Mapes, & R. A. Davis (Eds.), Cephalopods—Present and past: New insights and fresh perspectives (pp. 181–204). New York: Springer.
Raup, D. M. (1973). Depth inferences from vertically imbedded cephalopods. Lethaia, 6, 217–226.
Reyment, R. A. (1970). Vertically inbedded cephalopod shells. Some factors in the Distribution of fossil Cephalopods, 2. Palaeogeography, Palaeoclimatology, Palaeoecology, 7, 103–111.
Rickard, L. V. (1952). The Middle Devonian Cherry Valley Limestone of eastern New York. American Journal of Science, 250, 511–522.
Shindewolf, O. (1968). Analyse eines Ammoniten-Gehäuses. Akademie der Wissenschaften und der Literatur. Abhandlungen der Mathematisch-Naturwissenschaftlichen Klasse in Mainz, 8, 139–188.
Shoulga-Nesterenko, M. (1926). Nouvelles données sur l’organisation intérieure des conques des ammonites de l’étage d’Artinsk. Bulletin de la societé des Naturalistes de Moscou Section Géologique, 34, 81–99.
Tanabe, K., Fukuda, Y., & Obata, I. (1982). Formation and function of the siphuncle–septal neck structures on two Mesozoic ammonites. Transactions and Proceedings of the Paleontological Society of Japan, New Series, 128, 433–443.
Tanabe, K., & Landman, N. H. (1996). Septal neck-siphuncular complex of ammonoids. In N. H. Landman, K. Tanabe, & R. A. Davis (Eds.), Ammonoid paleobiology (pp. 129–165). New York: Plenum Press.
Tanabe, K., Landman, N. H., & Weitschat, W. (1993). Septal necks in Mesozoic Ammonoidea: Structure, ontogenetic development and evolution. In M. R. House (Ed.), The Ammonoidea: Environment, evolution and evolutionary change. Systematic Association Special (Vol. 47, pp. 57–84). Oxford: Clarenden Press.
Teichert, C. (1964). Morphology of the hard parts. In R. C. Moore (Ed.), Treatise on invertebrate paleontology, Part K, Mollusca 3 (pp. K1–K59). Lawrence, KS: Geological Society of America and University of Kansas Press.
Van Dyk, B. (1988). Love affair with a quarry. The Conservationist, 43(1), 46–48.
Wani, R., & Ikeda, H. (2006). Planispiral cephalopod shells as a sensitive indicator of modern and ancient bottom currents: New data from flow experiments with modern Nautilus pompilius. Palaios, 21, 289–297.
Weitschat, W., & Bandel, K. (1991). Organic components in phragmocones of boreal Triassic ammonoids: Implications for ammonoid biology. Paläontologische Zeitschrift, 65, 269–303.
Whiteley, T. E., & Kloc, G. L. (1999). Fossil lagerstätten of New York II. The cephalopods of the Cherry Valley Limestone. American Paleontologist, 7(2), 2–3.
Acknowledgments
The authors would like to thank the following: Paleontological Research Institute, Ithaca, New York, and Stephen Thurston, Morgan Hill, and Henry Towbin, all at the American Museum of Natural History. This research was supported by the Norman D. Newell Fund for Invertebrate Paleontology (AMNH) and the Lerner Gray Fund for Marine Research (AMNH).
Author information
Authors and Affiliations
Corresponding author
Additional information
Deceased: Susan M. Klofak.
Rights and permissions
About this article

Cite this article
Klofak, S.M., Landman, N.H. Ontogeny and morphology of the retrochoanitic septal necks of Agoniatites vanuxemi from the Middle Devonian Cherry Valley Limestone. Swiss J Palaeontol 134, 233–244 (2015). https://doi.org/10.1007/s13358-015-0092-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-015-0092-z