Special Issue: Cephalopods - Present and Past. Volume II
- Published:
Covariance of sutural complexity with whorl shape: evidence from intraspecific analyses of the Cretaceous ammonoid Desmoceras
Swiss Journal of Palaeontology volume 135, pages 1–10 (2016)
Abstract
In order to test the hypothesis that complex suture lines in ammonoids reinforced their shell strength, intraspecific variation of sutural complexity and shell morphologies (whorl shape and septal thickness) of the Cretaceous ammonoid Desmoceras latidorsatum (including three forms) from Madagascar were analyzed. The comparison between the sutural complexity and the whorl shape demonstrated that the more compressed or depressed forms have more complex suture lines than intermediate forms. The more compressed or depressed shells are generally expected to be weaker than intermediate ones that have almost circular whorl cross-sections. A comparison between sutural complexity balance (the ratio of the sutural complexities of the ventral and lateral parts) with the whorl shape demonstrated that the suture lines are more complex in the lateral area of the compressed shells and in the ventral area of the depressed shells. These relationships suggest that the suture lines were more complex in relatively more flattened parts of phragmocones, supporting that the complex suture reinforced the phragmocone against hydrostatic pressure.
Introduction
Ammonoids are a group of extinct cephalopods that possessed septate shells. The intersection between the outer shell wall and the septum is called “suture line”. The suture lines are generally more complex in geologically younger taxa than geologically older ones (e.g., Arkell 1957; Hirano et al. 1990; Korn 1992; Saunders and Work 1997; Saunders et al. 1999). This evolutionary trend in suture lines is used as a tool for classification of ammonoids (e.g., Arkell 1957; Miller et al. 1957; Kullmann and Wiedmann 1970; Korn et al. 2003, 2011; Monnet et al. 2011).
Several explanations have been proposed for the mechanism of sutural formation and for the physiological functions of suture lines in ammonoids. Explanations for the mechanism of sutural formation are the tie-point model (Seilacher 1975; Westermann 1975), the viscous fingering model (García-Ruiz et al. 1990; Checa and García-Ruiz 1996), and the reaction–diffusion model (Hammer 1999). Explanations for the physiological functions of suture lines are the reinforcing theory (Buckland 1836), concerning the transport of cameral liquid to the siphuncle (Mutvei 1967), and the efficiency of body attachment (Henderson 1984). Buckland (1836) hypothesized that the complex suture lines reinforced the outer shell against the hydrostatic pressure based on the observation of sutural pattern and whorl shape. This theory has been supported by many later studies (e.g., Westermann 1971, 1975; Ward 1980; Jacobs 1990; Hewitt and Westermann 1997; Hassan et al. 2002; De Blasio 2008). Jacobs (1990) analyzed the sutural distance of the succeeding septa and circumferential curvature (radius of curvature) of outer shell walls in Cretaceous baculitid ammonoids, and concluded that the vaults of septa contributed to reduce the tensional stress and maintained the outer shell during compression when hydrostatic load induced bending moments. The theoretical calculations and simulations in De Blasio (2008) suggested that complex septa reduced strain and stress in the phragmocone.
Alternatively, several studies did not support the reinforcing theory by Buckland (1836). Saunders (1995) demonstrated that the sutural complexity of the Paleozoic ammonoids did not correlate with the outer shell thickness, septal thickness and whorl shapes. Daniel et al. (1997) showed that fluted septa decreased shell strength using a computer model. Olóriz and Palmqvist (1995) demonstrated that the sutural complexity did not correlate with the estimated habitat depth in Mesozoic ammonoids. Besides, the covariations of shell morphologies in ammonoids (known as Buckman’s law of covariation) are explained by morphogenetic (Guex et al. 2003) or constructional constraints (Checa and García-Ruiz 1996; Monnet et al. 2011) rather than the physiological functions.
These previous studies mainly focused on the interspecific analyses (Westermann 1971, 1975; Ward 1980; Olóriz and Palmqvist 1995; Saunders 1995; Hewitt and Westermann 1997). However, intraspecific analyses using a large number of specimens have been rarely used to assess functional aspects of sutural complexity until now. Therefore, in this paper, the intraspecific correlations between sutural complexity and shell morphologies in several specimens of the Cretaceous ammonoid Desmoceras latidorsatum in various ontogenetic stages were analyzed. Similar to earlier studies (Buckland 1836; Seilacher 1988; Jacobs 1990; Saunders 1995), we focused on the relationships between streamlining and sutural complexity. In addition, comparisons between septal thickness and sutural complexity were carried out in this paper, similar to Saunders (1995).
Materials
In this study, 105 specimens of Desmoceras latidorsatum (Desmoceratidae, Ammonitina) from the Mahajanga area, Madagascar, were used (Fig. 1). Because of their good preservation, the suture line and shell morphologies could be measured easily. Although the exact fossil horizon is uncertain, the preservation states of all the examined specimens are quite similar, suggesting that the specimens are obtained from one locality or a single bed. The geological age of the examined specimens is assigned to the Albian according to Collignon (1963).

Photograph of Desmoceras latidorsatum from Madagascar. a, b Lateral and ventral views of D. latidorsatum forma complanata; c, d lateral and ventral views of D. latidorsatum forma media; e, f lateral and ventral views of D. latidorsatum forma inflata
Desmoceras latidorsatum shows wide intraspecific variations in shell morphology (Jacob 1908; Breistroffer 1933; Wiedmann and Dieni 1968; Cooper and Kennedy 1979; Kawabe and Haggart 2003; Kennedy and Klinger 2013). Among the examined 105 specimens, three forms can be distinguished based on shell depressions and constrictions (Fig. 1). According to Kennedy and Klinger (2013), these three forms were assigned to (1) forma complanata (19 specimens; Fig. 1a, b), with compressed shells and no or very weak constrictions, which corresponds to D. latidorsatum var. complanata (Jacob 1908); (2) forma media (79 specimens; Fig. 1c, d), with intermediate shells and 9–10 constrictions per whorl, which corresponds to D. latidorsatum var. media (Jacob 1908); and (3) forma inflata (seven specimens; Fig. 1e, f), with depressed shells and 6–7 constrictions per whorl, which corresponds to D. latidorsatum var. inflata (Breistroffer 1933). The term “forma” in this paper means simply variants, and does not mean pathologic forms. All specimens are deposited in the Mikasa City Museum, Hokkaido.
Methods
All specimens were polished along the median plane by means of silicon carbide powders (Fig. 2). The whorl shape, suture line, and the septal thickness were measured at four different shell diameters (15, 20, 25, and 30 mm). Shell diameter (D), whorl breadth (B), and whorl height (H) were measured using a caliper (Fig. 2a). Because the whorl breadth was measured after polishing the specimens along the median plane, the measured whorl breadth equals B/2. The measured whorl can simply be doubled to obtain the actual whorl breadth (B). The suture lines were examined from the venter to the umbilical seam for each diameter (Fig. 2b), although some suture lines cannot be studied due to the poor preservation. The suture lines were transcribed to paper by rotating the specimens little by little, using an optical microscope camera lucida. The suture lines were then digitalized with a digital scanner. In the next step, the length of the suture lines was measured. Septal thicknesses were measured on the polished surface using a digital optical microscope. Here, the septal thicknesses were measured at two points (Fig. 2c, d). These are (1) near the center of the septum, which approximates the maximum septal thickness, and (2) near the septal neck, which approximates the minimum septal thickness.

Measured shell morphology. a Phragmocone from ventral view; b traced suture line; c specimen polished along the median plane; d measured positions of septal thickness (near the center of septum and near the septal neck)
To analyze relationships among shell morphologies, four items are defined as below.
-
The whorl breadth/whorl height ratio (B/H) was used as an indicator for streamlining. It was calculated by whorl breadth (B) and whorl height (H). B/H is easy to measure and calculate, thus reducing measurement or other errors.
-
Index of sutural complexity (ISC) is the ratio of the curve length (CL) and whorl circumference (WC) (Fig. 2b). This method is the same as that used by Westermann (1971) and Ward (1980). The sutural components (the number of lobes and saddles) were consistent in three forms of Desmoceras latidorsatum (Fig. 3), and thus, the sinuosity of the suture line and vertical extent is reflected in this index.
Fig. 3 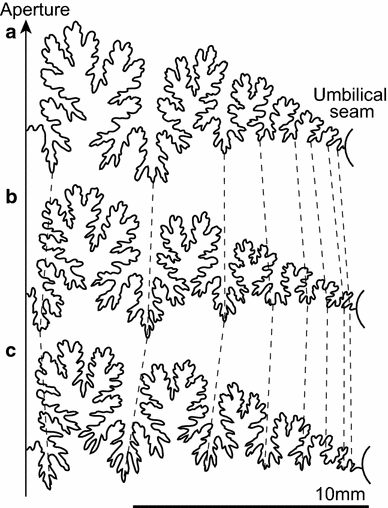
Suture lines of Desmoceras latidorsatum from Madagascar. a D. latidorsatum forma complanata, (DLC022, shell diameter of 25 mm); b D. latidorsatum forma media, (DLM111, shell diameter of 25 mm); c D. latidorsatum forma inflata, (DLI007, shell diameter of 25 mm)
-
The septal thickness near the center of the septum (STcenter) is defined as a percentage ratio of the original value of septal thickness near the center of the septum against shell diameter (D). This calculation was made in order to eliminate size variability.
-
The septal thickness near the septal neck (STneck) is defined as a percentage ratio of the original value of septal thickness near the septal neck against shell diameter (D). This calculation was made for the same purpose as definition of STcenter.

Assessment of measurement error
The suture lines are three dimensional on the inner surface of the phragmocone, so that the original suture shape must be distorted by transcribing it onto the two-dimensional paper. Therefore, the measurement error needs to be examined. Here, the measurement errors at shell diameters of 15 and 30 mm of two specimens of forma media (specimen nos. DLM021 and DLM091, respectively) were evaluated. The suture lines of each specimen at each diameter were transcribed five times (Table 1). The standard deviations of ISC for five times at the shell diameters of 15 and 30 mm were 0.12 and 0.08, and the mean ISC values with the 95 % confidence intervals were 7.94 ± 0.10 and 12.24 ± 0.07, respectively. The observed range of ISC values of forma media for the shell diameters of 15 and 30 mm was 6.84–10.02 (N = 37) and 8.68–12.30 (N = 38), respectively (Table 2), suggesting that the measurement error is negligible for the present analyses.
Results
The results of the measurements are summarized in Table 2 (see also Supplementary Material 1–4). ISC values increased with shell diameters in all forms. B/H values were almost constant through the ontogeny in forma media and decreased with shell diameters in forma complanata and forma inflata.
The correlations between ISC and other shell measurements (B/H, STcenter and STneck) of each shell diameter are summarized in Table 3. The comparisons between ISC and STcenter revealed no significant correlation in all forms (p > 0.05). The comparisons between ISC and STneck showed negative correlations in 20-mm shell diameter in forma media and in 15-mm shell diameter in forma inflata (p < 0.05).
ISC and B/H are positively correlated in all shell diameters in forma inflata and negatively correlated in 20–30-mm shell diameters in forma complanata (p < 0.05). In forma media, there were no significant correlations between ISC and B/H, through ontogeny (p > 0.05). The scatter diagrams of forma media (Fig. 4) showed distributions that tended to form inverse triangles: the more compressed and depressed shells of forma media have more complex suture lines, and the intermediate shells (1.00–1.10 of B/H) have suture lines with a wider range of ISC, especially in the later ontogenetic stages (20–30 mm in shell diameters, Fig. 4b–d). The positions of vertices of the inverted triangle were situated at 1.00–1.10 of B/H values, although the positions were slightly different, depending on the ontogenetic stages.

Scatter diagrams between ratio of whorl breadth and height (B/H) and index of sutural complexity (ISC). a Shell diameter of 15 mm; b shell diameter of 20 mm; c shell diameter of 25 mm; d shell diameter of 30 mm. The approximated reduced major axes are shown in the case of statistically significant correlation coefficients (p < 0.05)
Discussion
Sutural complexity in Desmoceras latidorsatum
The results of ISC analysis did not reveal a significant correlation with STcenter and STneck in almost all shell diameters in all forms (Table 3). These results suggest that sutural complexity may not be closely related to the septal thickness. The results also showed that sutural complexity correlates with whorl shape (Table 3; Fig. 4). According to theoretical and experimental analyses, a nearly circular tube is more resistant to the external hydrostatic pressure than a strongly oval tube (Estefen 1999). Therefore, the more compressed or depressed shells of ammonoids are possibly less resistant than the intermediate shells (i.e., 1.00 of B/H) against hydrostatic pressure. Accordingly, more complex suture lines are thought to reinforce the weaker shell whorls, because the more depressed or compressed shells are, the suture lines become more complex (Fig. 4). The reason why the vertices of inverse triangle of forma media are not exactly at 1.00 in B/H (Fig. 4) might be because the whorl shape is not perfectly circular in cross section (Fig. 2a).
Saunders (1995), Saunders and Work (1997) and this study suggest that the relationships between sutural complexity, whorl shape, and septal thickness are not consistent. According to Saunders (1995), the sutural complexity of Middle Devonian–Upper Permian ammonoids (agoniatitids, goniatitids, prolecanitids, and ceratitids) does not correlate with whorl shape. By contrast, Saunders and Work (1997) demonstrated that the sutural complexity of Lower Carboniferous–Lower Triassic prolecanitids correlated with whorl shape. These different relationships might relate to the difference in sutural components (number and complexity of lobes and saddles), lineages, and/or geological age.
Spatial factor to sutural complexity
It has been suggested that whorl width affects sutural complexity (the viscous fingering model; Checa and García-Ruiz 1996). The more compressed shells generally have a relatively wider space between the ventral wall of the outermost whorls and those of the previous whorl, and a relatively narrower space in the lateral area. Conversely, the more depressed shells generally have a relatively wider space in the lateral part and a relatively narrower space ventrally. Therefore, the tendency that the more compressed or depressed shells have more complex suture lines (Fig. 4; Table 3) is possibly derived from the relatively wider space in lateral or ventral areas of phragmocone.
In a further step, the suture lines were subdivided into their ventral and lateral parts (Fig. 5). The ventral part is between the external and lateral lobes, and the lateral part is between the lateral and 4th umbilical lobes (Fig. 5b). The boundary between these parts approximately corresponds to where the curvature of the shell shape changes markedly (Fig. 5c). The index of sutural complexity of each part was calculated similarly to the ISC calculation (Fig. 2b). Based on these calculations of the sutural complexities, the ISC balance, which indicates whether the sutural complexities contribute to either ventral part or lateral part, was calculated as

The suture lines of ventral and lateral parts. a Photograph of outer shell whorl; b suture line; c cross section of shell whorl
The scatter diagrams between B/H and ISC balance (Fig. 6) showed positive correlations in all the examined shell diameters (p < 0.05), which indicates that (1) in the more compressed shells (i.e., forma complanata), the lateral suture lines tend to be more complex than those in the ventral part; and (2) in the more depressed shells (i.e., forma inflata), the suture lines in the ventral part tend to be more complex than those in the lateral part. In the viscous fingering model, the relationship between provided space and undulation size (corresponding to sutural lobe size) was observed (Checa and García-Ruiz 1996). In addition, the positive correlations with the shell breadth and the number and size of sutural components were found in some Paleozoic prolecanitids and goniatitids (Checa and García-Ruiz 1996). However, the covariance of B/H and ISC balance in this paper (i.e., the suture lines in the lateral part tend to be more complex than those in the ventral part in the more compressed shells) shows no match with those in Checa and García-Ruiz (1996). This contradiction suggests that the more complex suture lines did not depend on the relatively wider space in lateral or ventral areas of phragmocone. Therefore, the covariance of B/H with ISC balance should be explained by other factors than the spatial constraints.

Scatter diagrams between ratio of whorl breadth and whorl height (B/H) and ISC balance. a shell diameter of 15 mm; b shell diameter of 20 mm; c shell diameter of 25 mm; d shell diameter of 30 mm. The approximated reduced major axes are shown, because all the correlation coefficients are statistically significant (p < 0.05)
The ventral area of the depressed shells (shells with high values of B/H) generally is more flattened, which indicates that the circle approximated to the venter has a larger shell whorl radius (Fig. 1f). Similarly, the lateral area of the compressed shells (shells with low values of B/H) generally is more flattened (Fig. 1b). Therefore, the positive correlations between B/H and ISC balance indicate that the suture lines are more complex in each flattened part of the shell in both compressed and depressed shells. Monnet et al. (2011) described similar covariation of suture line with streamlining in the Devonian ammonoids. They suggested that sutural complexity was caused by morphogenesis rather than functional factor, because the evolutionary trends of increasing of number of lobes were induced by different shell characters (streamlining and acute on the venter) in two lineages. However, the number of lobes does not covary with whorl shape in all forms of Desmoceras latidorsatum (Fig. 3). Covariations of suture line with whorl shape in the Cretaceous ammonoid Desmoceras latidorsatum are different from those of Monnet et al. (2011). Therefore, the explanations for suture line formation in Monnet et al. (2011) do not concur with this study. On the other hand, Jacobs (1990) reported tendencies similar to this study in the Cretaceous heteromorphic baculitids from the USA, and concluded that the vaults of septa contributed to reduce the tensional stress induced by hydrostatic load. Vertical extent of suture line is reflected in indicator of sutural complexity in Jacobs (1990) and ISC in this study. The agreement of findings in present analyses and that in Jacobs (1990) supports our conclusion that the more complex suture lines reinforced the shells against the hydrostatic pressure of surrounding seawater, at least in the examined Cretaceous Desmoceras latidorsatum.
References
Arkell, W. J. (1957). Introduction to mesozoic ammonoidea. In R. C. Moore (Ed.), Treatise on invertebrate paleontology (Vol. L, pp. 81–129). Boulder, Lawrence: Geological Society of America, University of Kansas Press.
Breistroffer, M. (1933). Étude de l’étage Albien dans le massif de la Chartreuse (Isère et Savoie). Travaux du Laboratoire de Géologie de l’Université Grenoble, 17, 1–50.
Buckland, W. (1836). Geology and mineralogy considered with reference to natural theology. London: Pickering.
Checa, A. G., & García-Ruiz, J. M. (1996). Morphogenesis of the septum in ammonoids. In N. H. Landman, K. Tanabe, & R. A. Davis (Eds.), Ammonoid paleobiology (pp. 253–296). New York: Prenum.
Collignon, M. (1963). Atlas des fossiles caractéristiques de Madagascar (Ammonites), Fascicule X (Albien). Tananarive: Service Géologique de Madagascar.
Cooper, M. R., & Kennedy, W. J. (1979). Uppermost Albian (Stoliczkaia dispar Zone) ammonites from the Angolan littoral. Annals of the South African Museum, 77(10), 175–308.
Daniel, T. L., Helmuth, B. S., Saunders, W. B., & Ward, P. D. (1997). Septal complexity in ammonoid cephalopods increased mechanical risk and limited depth. Paleobiology, 23(4), 470–481.
De Blasio, F. V. (2008). The role of suture complexity in diminishing strain and stress in ammonoid phragmocones. Lethaia, 41(1), 15–24.
Estefen, S. F. (1999). Collapse behaviour of intact and damaged deepwater pipelines and the influence of the reeling method of installation. Journal of Constructional Steel Research, 50(2), 99–114.
García-Ruiz, J. M., Checa, A., & Rivas, P. (1990). On the origin of ammonite sutures. Paleobiology, 16(3), 349–354.
Guex, J., Koch, A., O’Dogherty, L., & Bucher, H. (2003). A morphogenetic explanation of Buckman’s law of covariation. Bulletin de la Société Géologique de France, 174(6), 603–606.
Hammer, Ø. (1999). The development of ammonoid septa: An epithelial invagination process controlled by morphogens? Historical Biology, 13(2–3), 153–171.
Hassan, M. A., Westermann, G. E. G., Hewitt, R. A., & Dokainish, M. A. (2002). Finite-element analysis of simulated ammonoid septa (extinct Cephalopoda): septal and sutural complexities do not reduce strength. Paleobiology, 28(1), 113–126.
Henderson, R. A. (1984). A muscle attachment proposal for septal function in Mesozoic ammonites. Palaeontology, 27(3), 461–486.
Hewitt, R. A., & Westermann, G. E. G. (1997). Mechanical significance of ammonoid septa with complex sutures. Lethaia, 30(3), 205–212.
Hirano, H., Okamoto, T., & Hattori, K. (1990). Evolution of some Late Cretaceous desmoceratine ammonoids. Transactions and Proceedings of the Palaeontological Society of Japan, New Series, 157, 382–411.
Jacob, C. (1908). Étude sur quelques Ammonites du Crétacé moyen. Mémoires de la Société Géologique de France, Paléontologie, 15, Mémoire, 38(1907), 1–64.
Jacobs, D. (1990). Sutural patterns and shell stress in Baculites with implications for other cephalopod shell morphologies. Paleobiology, 16(3), 336–348.
Kawabe, F., & Haggart, J. W. (2003). The ammonoid Desmoceras in the Upper Albian (Lower Cretaceous) of Japan. Journal of Paleontology, 77(2), 314–322.
Kennedy, W. J., & Klinger, H. C. (2013). Cretaceous faunas from Zululand and Natal, South Africa. The ammonite Subfamily Desmoceratinae Zittel, 1895. African Natural History, 9, 39–54.
Korn, D. (1992). Relationships between shell form, septal construction and suture line in clymeniid cephalopods (Ammonoidea; Upper Devonian). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 185, 115–130.
Korn, D., Bockwinkel, J., Ebbighausen, V., & Walton, S. A. (2011). Beloceras, the most multilobate Late Devonian ammonoid. Bulletin of Geosciences, 86(1), 1–28.
Korn, D., Ebbighausen, V., Bockwinkel, J., & Klug, C. (2003). The A-mode sutural ontogeny in prolecanitid ammonoids. Palaeontology, 46(6), 1123–1132.
Kullmann, J., & Wiedmann, J. (1970). Significance of sutures in phylogeny of Ammonoidea. The University of Kansas, Paleontological Contributions, 47, 1–32.
Miller, A. K., Furnish, W. M., & Schindewolf, O. H. (1957). Paleozoic ammonoidea. In R. C. Moore (Ed.), Treatise on invertebrate paleontology (Vol. L, pp. 11–79). Boulder, Lawrence: Geological Society of America, University of Kansas Press.
Monnet, C., Klug, C., & De Baets, K. (2011). Parallel evolution controlled by adaptation and covariation in ammonoid cephalopods. BMC Evolutionary Biology, 11(115), 1–21.
Mutvei, H. (1967). On the microscopic shell structure in some Jurassic ammonoids. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 129, 157–166.
Olóriz, F., & Palmqvist, P. (1995). Sutural complexity and bathymetry in ammonites: Fact or artifact? Lethaia, 28(2), 167–170.
Saunders, W. B. (1995). The ammonoid suture problem: Relationships between shell and septum thickness and suture complexity in Paleozoic ammonoids. Paleobiology, 21(3), 343–355.
Saunders, W. B., & Work, D. M. (1997). Evolution of shell morphology and suture complexity in Paleozoic prolecanitids, the rootstock of Mesozoic ammonoids. Paleobiology, 23(3), 301–325.
Saunders, W. B., Work, D. M., & Nikolaeva, S. V. (1999). Evolution of complexity in Paleozoic ammonoid sutures. Science, 286, 760–763.
Seilacher, A. (1975). Mechanische Simulation und funktionelle Evolution des Ammoniten-Septums. Paläontologische Zeitschrift, 49(3), 268–286.
Seilacher, A. (1988). Why are nautiloid and ammonite sutures so different? Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 177, 41–69.
Ward, P. D. (1980). Comparative shell shape distribution in Jurassic-Cretaceous ammonites and Jurassic-Tertiary nautilids. Paleobiology, 6(1), 32–43.
Westermann, G. E. G. (1971). Form, structure and function of shell and siphuncle in coiled Mesozoic ammonoids. Royal Ontario Museum, Life Sciences Contributions, 78, 1–39.
Westermann, G. E. G. (1975). Model for origin, function, and fabrication of fluted cephalopod septa. Paläontologische Zeitschrift, 49(3), 235–253.
Wiedmann, J., & Dieni, I. (1968). Die Kreide Sardiniens und ihre Cephalopoden. Palaeontographia Italica, 64, 1–171.
Acknowledgments
We are grateful to R. Majima (Yokohama National University), T. Ubukata (Kyoto University), R. G. Jenkins (Kanazawa University), K. Kurihara (Hokkaido Museum), L. Doguzhaeva (Swedish Museum of Natural History), M. Utsunomiya (Geological Survey of Japan), K. Noshita (University of Tokyo), T. Toyofuku (JAMSTEC), S. Sakano (Waseda University), and T. Hosono (Waseda University), K. Ikuno (Yokohama National University) for their fruitful suggestions, Y. Takeda (University of Tokyo) for collecting literatures, and C. Klug (Universität Zürich) and two anonymous reviewers for their critical comments on an early draft. This study was supported by Grant-in-Aid for Young Scientists (Nos. 21740369 and 26800264 to RW), the Japan Science and Technology Agency (JST; to RW), and a grant from Faculty of Environment and Information Sciences, Yokohama National University (to RW).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Aiba, D., Wani, R. Covariance of sutural complexity with whorl shape: evidence from intraspecific analyses of the Cretaceous ammonoid Desmoceras . Swiss J Palaeontol 135, 1–10 (2016). https://doi.org/10.1007/s13358-015-0103-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-015-0103-0